Institut pour une
triarticulation sociale
|
||||||||||||||||||
| Accueil
nv>actualités
|
|
|||||||||||||||||
Trad. v.01, F. Germani
au 12/02/2022
sur la base d'une traduction automatique dont je n'ai pu vérifier que la cohérence globale sans pour autant avoir les connaissances médicales nécessaires au choix des termes. Aux lecteurs plus qualifiés de me signaler les améliorations possibles. Préambule sur Gerhard Kienle par Wolfgang Schad L'essai suivant ("Tübingen 1950. Reproduit sous forme de manuscrit") n'a jamais été publié par PD, Dr. med. habil. Gerhard Kienle n'a jamais été pensé pour la publication. Il l'a écrit à l'âge de 27 ans comme une ébauche crue destinée à stimuler la discussion entre les médecins familiarisés avec les questions de Rudolf Steiner sur la physiologie nerveuse. Il ne l'aurait jamais publiée, car il était parfaitement conscient du caractère inachevé et immature de ce document. Par la présente édition, l'auteur lui-même ne devait et ne doit jamais devenir l'objet de la critique, car Gerhard Kienle ne se distingue pas de lui-même en tant qu'auteur ou éditeur. Une citation critique ne peut donc être faite qu'en mentionnant ces circonstances spéciales.Tout au plus peut-on critiquer la décision de l'éditeur de rendre l'ouvrage accessible au public. Ce qui justifie cette démarche, c'est la richesse de la réflexion et l'intégration d'une littérature abondante, que les personnes intéressées pourront utiliser pour leur propre travail sur le sujet. En 1981, Gerhard Kienle avait encore l'intention de former lui-même un groupe de travail pour retravailler le sujet, afin d'obtenir pour la première fois un traitement compétent de celui-ci, ce qui n'a pas pu se faire en raison de son décès prématuré. La publication en deux volumes présentée ici a pour but de mettre à disposition les travaux préparatoires réalisés jusqu'à présent, ainsi que l'ouvrage suivant, comme condition préalable à tout traitement futur. Petite biographie de Gerhard Kienle par Peter Selg Sur Wikipédia : https://de-m-wikipedia-org.translate.goog/wiki/Gerhard_Kienle?_x_tr_sl=auto&_x_tr_tl=fr&_x_tr_hl=en ***
GERHARD KIENLELes questions fondamentales de la physiologie nerveuseIntroductionDans la vie scientifique actuelle, la question du rapport entre la corporéité de l’homme et son psycho-spirituel est traitée de la manière la plus diverse. Les concepts que l’on admet comme bases physiologiques sont le plus souvent tirées du monde de l’inorganique/anorganique, c’est-à-dire de la physique et de la chimie. Les représentations physiologiques datent presque toutes d’une époque où les plus grands efforts et tentatives ont été faits pour expliquer les phénomènes physiologiques de manière matérialiste. Les notions qui existent aujourd’hui sur le système nerveux seront ici examinées de manière critique afin de déterminer dans quelle mesure elles reposent réellement sur l’observation et dans quelle mesure elles contiennent des représentations hypothétiques. On se demande en particulier si les notions chimiques et physiques suffisent à elles seules à fonder des représentations physiologiques. On rompt cependant avec le principe qui consiste à chercher l’essence du vivant et de l’âme dans l’opacité des processus matériels compliqués. Cette attitude scientifique s’exprime déjà très clairement chez Lotze ; il écrit en 1842 dans le Handwörterbuch der Physiologie, vol. I, p. XXVI : « Ni le sang ni les nerfs ne sont proprement animés, mais la vie appartient au tout et est, strictement pris, un résumé/une saisie ensemble de processus inanimés/non parcourus de vie ». Les suggestions concernant ces questions sont dues à Rudolf Steiner, en particulier à son ouvrage Von Seelenrätseln. Les explications sont une tentative de répondre aux questions qui se posent sur les bases de la recherche expérimentale et de développer les concepts purement à partir de la chose. Ce faisant, on s’est efforcé de les former le plus indépendamment possible des précédentes/de celles jusque là. Ce traité veut être considéré comme une compilation provisoire de faits pour la formation d’un jugement sur les questions correspondantes. L’aspect scientifique peut également revêtir un caractère provisoire, car il est impossible de connaître tous les travaux et recherches correspondants et de pouvoir juger pleinement de la fiabilité de ceux qui ont été découverts. C’est pourquoi, dans un premier temps, seuls les points de vue seront soumis à la discussion. Ce n’est qu’à l’issue de cette discussion que des détails contraignants pourront être établis. La situation du problème peut être résumée de la manière suivante : 1. l’homme se présente comme un fait matériel, au même titre que la nature organique et inorganique. Mais il est en même temps vécu comme un être psychique et spirituel. Les phénomènes psychologiques peuvent-ils être expliqués par des processus matériels dans l’organisme ? Dans ce cas, l’homme pourrait être appréhendé et compris de la même manière que la matière inorganique, il ne serait qu’un cas particulier des processus généraux de la nature. Les réactions de l’organisme, en particulier lorsqu’elles se manifestent par des mouvements, peuvent avoir pour cause observable des stimulations des organes sensoriels. Pour une explication matérielle, les voies de conduction entre les organes sensoriels et moteurs doivent transmettre les changements d’état matériels en tant qu’intermédiaires de cause à effet. Les nerfs sont hypothétiquement abordés comme de tels câbles pour la transmission de l’excitation et le système nerveux central comme le centre de commutation correspondant. Il s’agit d’examiner si un tel mode d’explication est possible et si les nerfs possèdent effectivement la fonction qui leur est attribuée. 2. Pour traiter un tel problème, il n’est pas possible d’entrer dans des spéculations ou des postulats de philosophie naturelle : en particulier, l’exigence selon laquelle, lors du traitement de questions physiologiques, même si elles ne peuvent pas être expliquées physico-chimiquement, on doit se comporter comme si cela était possible, doit être rejetée comme étant en contradiction avec la méthodologie scientifique. La formation d’un jugement ne peut être invoquée que sur la base de faits observables. Les études méthodologiques servent à déterminer ce que les différentes observations révèlent de la problématique en question. Il a été constaté que la stimulation artificielle des nerfs ne peut rien révéler de leur fonctionnement normal. 3. La fonction sensorielle n’est pas la conduction hypothétique d’un stimulus. L’homme et l’animal ne réagissent que très rarement à des stimuli simples, mais le plus souvent à des objets, des formes ou des situations. La perception ne comprend pas seulement le stimulus sensoriel, mais aussi un élément idéel appelé représentation. Celui-ci ne peut pas être expliqué de manière sommative, mais vient s’ajouter en tant qu’élément immatériel. Les seuils de stimulation des organes sensoriels sont déterminés par l’objet qu’ils perçoivent. Le processus de perception ne peut pas être considéré comme une image analogue à l’image optique dans l’œil. Sa constitution implique une activité et un mouvement internes et externes. Les principes de la conduction expérimentale ne peuvent pas être confirmés dans des conditions/rapports physiologiques. 4. Le mouvement n’est pas, d’un point de vue fonctionnel, le résultat de l’action d’organes isolés, il se présente dans sa configuration comme quelque chose d’indépendant en soi et ordonne les organes en soi. L’activité sensorielle et la perception font partie de sa constitution de différentes manières, et pas seulement pour la stimulation. 5. L’activité motrice doit être différenciée en elle-même. Tout d’abord, c’est le déploiement de la force qui ressort en tant que tel, ensuite le mouvement se présente comme une forme et est soumis aux principes de la forme. La forme elle-même peut être impliquée dans différents contextes, de l’intérieur en tant qu’expression de contenus psychiques, vers l’extérieur elle est mise en relation avec des actions par l’intention de la volonté et se présente comme une prestation. 6. La perception et le mouvement ne sont pas des fonctions séparées et ne peuvent pas non plus être classés dans des théories de conduction. Ils rendent aussi nécessaire un nouveau concept de l’espace et du temps. Contrairement au caractère discontinu de la physique, l’espace et le temps sont ici soumis à la continuité. Les deux peuvent se fondre l’un dans l’autre, des processus passés peuvent contribuer à déterminer les processus présents et des processus apparaissant plus tard dans le temps peuvent interférer avec des processus antérieurs. Le phénomène de la conduction apparaît comme lié aux conditions expérimentales et est un produit artificiel pour les conditions physiologiques. 7. Des réflexes, pour l’apparition desquels on a rendu responsables certaines voies ou conditions structurelles, se sont avérés être des faits physiologiques lors d’un examen plus approfondi. 8. Les prémisses d’une théorie de la conduction s’étant révélées inexactes/non pertinentes, on a procédé à une révision du concept de nerf et des représentations de l’interaction entre le psychique-spirituel et le corporel. 9. La perception commence dans l’organe sensoriel lui-même. Le système nerveux est le support de l’activité de représentation. Les localisations et les perceptions individuelles sont des prestations de représentation. Les combinaisons et les constructions ne peuvent être effectuées que secondairement, une fois la ponctualité atteinte par la différenciation. Les éléments des représentations peuvent être saisis en tant qu’entités uniformes avec leur propre dynamique de force, ce que l’on appelle les préformes. En devenant une représentation, ils perdent toute activité propre et deviennent une pure image. Ceci est provoqué par la fonction du nerf. Le cerveau lui-même est un organe miroir, la localisation est le résultat de la mémorisation des fonctions. 10. Dans le cas de la mémoire, la formation d’engrammes est un processus parallèle à la représentation. Celle-ci ne se produit pas dans la substance cérébrale, mais dans le métabolisme. À chaque acte de mémoire, de nouvelles représentations sont formées par la perception de l’engramme. 11. l’activité de la volonté intervient directement dans le métabolisme. Elle ne dépend du système nerveux que dans la mesure où elle est stimulée par l’activité sensorielle et s’oriente en fonction des perceptions. 12. Les nerfs dits sensitifs transmettent la perception du monde extérieur ou des processus internes, dans la mesure où ils sont devenus des faits matériels. 13. Les nerfs dits moteurs transmettent les processus internes dans la mesure où ils se tournent vers l’organisme. Ce n’est que par la perception interne des organes qu’ils peuvent être trouvés par la volonté, de même que le déploiement de force et le mouvement sont ressentis par eux, mais ne sont perçus pleinement consciemment que par l’image de la perception externe. 14. il faut distinguer les différents niveaux de conscience qui coexistent, se succèdent et s’imbriquent les uns dans les autres. Selon le mode de formation des représentations, il faut distinguer phylogénétiquement et ontogénétiquement la conscience de l’image et la conscience de l’objet. La seconde apparaît toujours en même temps que des pédiments de la première. 15. L’activité sensorielle stimule certes l’ensemble de la vie psychique et peut même avoir un effet suggestif, mais les véritables impulsions se trouvent dans la volonté, les représentations ne font qu’orienter, elles ne déterminent pas. Si elles ont un caractère de préfiguration, elles peuvent avoir un effet affectif. La liberté est rendue physiologiquement possible par la libre disposition des représentations qui, en tant que pures images, n’agissent pas sur la volonté ; ce n’est que lorsque les représentations sont à nouveau vivifiées par une activité propre qu’elles peuvent impulser la volonté et inverser la direction animale du processus en une humaine. 16. Il n’est pas possible d’élaborer une théorie uniforme sur la relation corps-esprit, les rapports sont différenciés et doivent être étudiés spécifiquement pour chaque question. C’est pourquoi presque toutes les théories peuvent être prouvées et réfutées sur la base de faits individuels/particuliers. Les théories doivent être remplacées par des observations individuelles/particulières concrètes. L’organisme humain n’est pas un cas particulier des processus généraux de la nature, mais en lui, le matériel et le spirituel interagissent de manière différenciée, c’est pourquoi d’autres concepts que ceux utilisés dans les sciences naturelles inorganiques sont nécessaires pour comprendre les faits. Les difficultés méthodologiques qui en découlent ne peuvent pas être évitées/contournées. Les questions fondamentales de la physiologie nerveuse La physiologie nerveuse a apporté un matériel
factuel extraordinairement important grâce à des études
cliniques et expérimentales, accompagnées d’observations
anatomiques et histologiques variées correspondantes. Jusqu’à
présent, les résultats ont défié toutes les tentatives de les
classer sans contradiction dans les systèmes et théories
connus. Étant donné que les conceptions relatives à la
fonction du système nerveux constituent un point clé de toute
la physiologie et de l’anthropologie, il est impossible de
faire l’impasse sur la problématique qui se pose ici, si l’on
ne veut pas se soumettre à des approches dogmatiques qui se
justifient par le caractère inexpliqué des problèmes. Comme il est tout à fait possible que nos
idées actuelles contiennent des hypothèses qui ont été
considérées comme utilisables à des stades antérieurs de la
recherche, mais qui ont été utilisées par la suite comme des
faits sans être vérifiées, malgré des conditions différentes,
il est nécessaire d’aborder le phénomène initial et sa
problématique au stade précédant la formation des concepts
scientifiques proprement dits. Ensuite, les conditions
méthodologiques pour l’interprétation des résultats
d’expériences doivent être discutées. Les différentes
questions pourront alors être examinées plus en détail à
l’aide des principes possibles d’explication. Le sécrétoire, dans la mesure où il se
présente comme vie intérieure, peut directement seulement être
vécu dans le propre horizon de la conscience. Pour l’autre
homme et pour l’animal, dans la mesure où il s’exprime, on
dépend de l’observation indirecte, l’animal étant limité par
le manque de langage. Dans la mesure où l’on analyse l’organisme dans ses parties, il perd sa capacité d’expression pour l’âme. C’est par les yeux, par exemple, que l’on peut avoir l’impression la plus forte de la présence personnelle d’un autre être humain ; cela disparaît complètement lorsqu’on examine le fond de l’œil ou les détails de l’iris. De même, on ne peut pas voir le contexte d’une action dans la contraction d’une fibrille musculaire. Plus on pénètre analytiquement dans les organes, plus il devient impossible de vivre la « totalité de l’âme » : celle-ci ne peut alors être qu’interprétée. L’affirmation selon laquelle on ne peut rien savoir de la vie intérieure d’autrui, que celle-ci ne peut être découverte et interprétée que sur la base des réactions des organes, ne vaut que pour la situation de comportement analytique extrême. Voilà pour la situation psychologique de départ. Si l’on applique maintenant l’observation de la nature à l’homme, le premier problème est le suivant : comment l’âme interagit-elle avec les organes ? De manière tangible, on n’a devant soi que des organes matériels, et si on les aborde avec des outils chimiques et physiques, on ne peut bien sûr rien trouver de psychique. Le calcaire dans l’homme est tout d’abord chimiquement identique au calcaire dans le reste de la nature. L’organisme peut-il être compris, selon les lois chimiques et physiques, comme une interaction des données matérielles à la manière des processus naturels inorganiques ? Si l’on considère l’anatomie, les perceptions sont transmises par les organes sensoriels et les mouvements sont provoqués par la musculature. Ces deux groupes d’organes matériellement saisissables sont-ils aussi reliés par des effets matériels, de la même manière que les processus naturels inorganiques sont reliés entre eux par des forces physiques ? Or, les deux types d’organes sont reliés par deux systèmes matériels, le système sanguin et le système nerveux. Si l’on coupe le nerf menant à un organe sensoriel, la perception sensorielle s’éteint, la coupure du nerf moteur menant au muscle produit l’immobilité. Il en va de même pour le sang. Une stimulation artificielle, qui n’a rien de commun avec les processus normaux, peut provoquer une impression sensorielle dans le nerf sensoriel et une contraction musculaire dans le nerf moteur. Une telle expérience correspond tout à fait à la méthode de travail de science de la nature. On observe un processus, par exemple la congélation de l’eau, et on examine maintenant ce qui peut le provoquer et l’influencer. Ensuite, on avance pas à pas. La même méthode est appliquée au muscle. Le mouvement repose sur la contraction des muscles, on crée donc les conditions les plus diverses dans lesquelles une contraction musculaire peut avoir lieu. L’une d’entre elles est justement la stimulation du nerf d’alimentation, et c’est aussi la seule que l’on puisse imaginer dans un premier temps dans des conditions physiologiques. Les résultats décrits précédemment confirment cette hypothèse. La transmission d’une excitation matérielle
dans l’organe des sens par le système nerveux jusqu’à l’organe
du succès permettrait d’expliquer l’organisme humain et animal
selon les principes de l’observation inorganique de la nature,
en se basant sur la géométrie spatiale des connexions
nerveuses. On pourrait se passer, pour la formation de
théories physiologiques, d’un élément psychique particulier,
non directement observable par les sens. Ce qui apparaît à
l’âme ne serait alors que l’expression de processus matériels
et s’expliquerait entièrement par le système nerveux. 1. comme le psychique n’est observable qu’indirectement chez l’homme et l’animal, les conditions de l’observation psychologique doivent être claires si l’on veut dire quelque chose sur le rapport entre le psychique et un organe. Les interventions matérielles dans l’organisme ne peuvent donc pas être prises en compte pour le mode d’action du psychisme. 3. Comme c’est tout l’organisme qui s’avère être l’organe d’expression de l’âme, l’âme ne peut pas être localisée a priori sur un organe, comme le cerveau par exemple. Les tentatives d’interventions chirurgicales ne permettent que des conclusions psychologiques sur l’âme sans la partie de l’organe qui a été enlevée, mais on ne peut jamais constater que l’âme n’agit pas (1). Parallélisme psychophysique Pour penser que l’organe de réception et l’organe de réussite sont mécaniquement liés, il faut postuler une voie de conduction hypothétique qui transmet les excitations à l’instar d’une mèche ou de câbles électriques. De telles ondes d’excitation peuvent être représentées expérimentalement dans le système nerveux. Comme on peut également observer dans l’activité physiologique des changements d’état qui ressemblent à des excitations à variation rapide, les nerfs sensitifs sont interprétés comme la partie ascendante de cette voie de conduction hypothétique et les nerfs nécessaires à la motricité et conduisant aux organes de la réussite comme des voies descendantes. Selon cette conception, la répartition des excitations afférentes sur les voies efférentes est réglée par le système nerveux central. La théorie des réflexes qui en découle a d’abord postulé des voies et des connexions strictement isolées. La théorie des neurones, telle qu’elle a été développée par Forel, His, puis définitivement par Waldeyer et développée par R. y Cajal, en a fourni les bases anatomiques. L’archétype du réflexe est présenté dans le réflexe propre. Une excitation du fuseau musculaire retourne au muscle via deux autres neurones et son succès est strictement déterminé par la simple conduction. Les autres réflexes ne sont considérés que comme de simples/pures complications.Buytendijk (2) caractérise cette conception de la manière suivante : 1. les réflexes simples sont primaires, les contextes compliqués secondaires. 2. chaque réflexe a une trajectoire et un centre définis, il est donc strictement localisé. 3. toute activité est en fin de compte une réactivité. 4. les processus sensitifs et moteurs sont fondamentalement différents. 5. les processus sensibles sont la cause des processus moteurs, ces derniers sont donc clairement déterminés par les premiers. Ainsi, si a un stimulus n’est pas répondu par un seul effet (réflexe propre), mais qu’il provoque une activité prolongée, celle-ci est considérée comme commandée par un centre. En effet, on peut également trouver certaines localisations dans le système nerveux central. L’« apprentissage » est expliqué par la « mise en place » de nouvelles voies, on distingue donc les réflexes innés et les réflexes acquis conditionnels. Les différentes théories sont connues en détail, il n’est pas nécessaire de s’attarder sur la théorie des neurofibrilles, puisqu’il s’agit du même principe. Afin de ne pas devoir nier le côté subjectif de l’âme, la théorie du parallélisme psychophysique a été élaborée : Le psychique ne ferait donc qu’accompagner les processus physiques en parallèle (3). Avant d’entrer dans les détails de ces théories, il faut vérifier si les réactions ne se produisent vraiment qu’en réponse à des stimuli, si cette condition préalable est valable. La « loi des énergies sensorielles spécifiques » de Joh. Müller correspond à un tel principe de conduction mécaniste, selon lequel un organe sensoriel ou son nerf ne transmet toujours que la même impression sensorielle, indépendamment du stimulus. Que l’on stimule l’œil par la lumière, un coup, une pression, l’électricité ou autre, le résultat est toujours une sensation lumineuse. L’interprétation est ouverte pour savoir si cela donne une explication générale ou s’il s’agit seulement d’un phénomène spécial. Kries (4) n’admet pas ce principe sans réserve. Weizsäcker justifie abondamment sa réfutation (5). Tout d’abord, un organe sensoriel peut subir un changement de fonction, par exemple dans le cas pathologique, de plus la quantité du stimulus modifie tout à fait la qualité, par exemple le chatouillement et la pression. Le principe de conduction correspondait également à la conception selon laquelle seule l’image d’excitation serait représentée dans la conscience, conformément au processus de représentation optique dans l’œil. Les perceptions se composeraient des sensations individuelles et seraient conditionnées par l’état respectif des organes. Si l’on a des sensations isolées, c’est l’exception, on vit d’abord le contenu et les objets. Depuis Ehrenfels (6), la psychologie de la forme s’est longuement penchée sur la question de savoir si les perceptions ne sont que des sensations individuelles additives. Un nouveau principe non sensoriel doit s’ajouter aux impressions sensorielles pour que des objets en résultent. Ce principe supplémentaire n’est pas donné avec les sens et se trouve dans l’activité. Il dépend de notre propre activité. Lorsque l’on retire de la sensorialité ce principe qui s’y déploie et qu’on le met en relation avec d’autres, on le désigne alors comme concept ou idée (7). Cela est particulièrement bien observable lorsque des parties sont ambiguës. Les sommets d’un pentagone régulier peuvent être vus comme un cercle ou un pentagone. Il en va de même pour un groupe de figures dont la figure et le fond sont interchangeables, par exemple dans les bandes en relief ou dans le découpage qui représente un calice comme figure découpée et deux visages qui se regardent comme contours de base (8). Les fragments sont toujours classés en fonction de leur relation idéale avec un ensemble. Si l’on observe le plan d’un cube incliné d’une manière ou d’une autre, même s’il est borgne, on peut voir un dessin plan ou deux cubes qui se tiennent alternativement. L’impression de plan ou d’espace dépend également de l’activité personnelle. Dans quelle mesure l’inversion des deux corps possibles se fait de manière volontaire ou involontaire, c’est une question de discipline de sa propre activité de représentation. Il en va de même pour toutes les autres représentations spatiales, par exemple pour l’inversion de König, l’escalier de Thier, etc. (8a). L’imprégnation idéelle des sensations doit être appelée activité de représentation. La spatialité des perceptions repose également sur l’activité de représentation. Lorsque les yeux sont fermés, une surface grise apparaît devant les yeux, qui est avancée lors de la vision (cf. note 6, Katz). Les objets sont perçus dans un espace tridimensionnel, alors que la surface bidimensionnelle est suffisante pour les représentations purement picturales. Dans la perception normale du visage, elle apparaît comme fond. C’est pourquoi, dans le cas de figures avec possibilité d’échange de base, la figure envisagée sort du fond bidimensionnel et est vécue comme un objet — même plat — dans l’espace tridimensionnel. Les enfants aveugles de naissance opérés doivent d’abord acquérir l’espace (9), leur propre activité est nécessaire pour cela. Ainsi, toute spatialité dans la perception est déjà une performance de l’activité de représentation. Le regroupement idéel d’impressions individuelles en un tout se produit dans tous les domaines sensoriels. Les mélodies sont également une unité. Katz (6) fait remarquer que si l’on écoute la radio et qu’une émission est perturbée et carrément hachée, c’est-à-dire qu’aucun sens n’émerge de la simple juxtaposition d’impressions, on peut néanmoins entendre l’émission proprement dite. La spatialité ne peut d’abord être vécue passivement que sur son propre corps, mais elle n’est pas exclusivement une performance du sens du toucher ; si, en cas d’hémiplégie de Brown-Sequard, seule la sensibilité à la température est conservée, il est néanmoins possible de localiser (10). Pour palper des objets, un mouvement est nécessaire, le toucher pouvant également se faire indirectement avec un objet saisi. Une activité externe ou interne est toujours nécessaire. Les objets peuvent également être reconnus par des sensations vibratoires ; les aveugles (11) peuvent apprendre à le faire. Les poissons ont une sorte de « perception à distance » des objets via les organes latéraux (12). Les figures et les rapports de taille peuvent également être détectés de manière assez précise à partir du mouvement en tant que tel ; plus on le rend mécanique, par exemple en raidissant l’articulation du coude, moins l’estimation est précise (13). Mais certaines qualités de perception peuvent également être obtenues par le mouvement, comme les qualités tactiles rugueuses, lisses, élastiques, graisseuses, molles, dures, etc. (11). La perception n’est absolument pas toujours liée à un organe sensoriel. Si un organe est perturbé, la perception de l’objet peut aussi, dans une certaine mesure, se faire par un autre organe, si la capacité d’observation sur cet organe et la transformation entre les représentations de différents domaines sensoriels ont été exercées. Dans un premier temps, on peut écrire correctement avec la main gauche, les pieds ou la langue sans exercice particulier, les chiffres écrits sur la paume de la main sont reconnus sans problème. Pour plus de détails, voir Kamm (14) et Bethe (15). Ensuite, le développement de la capacité tactile est connu chez les aveugles. Les sensations vibratoires en particulier peuvent être développées de manière extraordinaire. En posant les mains sur le larynx de la personne qui parle, un sourd peut très bien comprendre la parole ; il peut aussi parfois bien suivre des événements dans l’environnement, comme la fermeture d’une porte, des grincements, etc. Helen Keller, qui dépendait exclusivement de son sens du toucher, a atteint des capacités tout à fait extraordinaires. Elle vivait tout son environnement presque comme une personne normale. La musique aussi peut être vécue de manière artistique (16). On connaît le cas de Sutermeister qui, après 55 ans de surdité, est devenu un passionné de musique (17) ; contrairement à Helen Keller, il percevait la musique et la parole à travers les sensations vibratoires de la cage thoracique, et ce par le dos. (Description détaillée des phénomènes chez les sourds-muets chez Gutzmann [18]). Goldstein décrit un cas de cécité de l’âme suite à une lésion cérébrale, dans lequel seul le point de fixation, la macula, permettait de voir correctement (19). Ce patient pouvait lire sans problème en suivant les lettres avec les mouvements des yeux, de même qu’il pouvait palper les contours des objets sans s’en rendre compte. Mais les représentations appelées figures ont aussi en elles une dynamique indépendante, comme on peut le voir dans les illusions d’optique connues. Dans le cas de figures ambiguës, l’illusion change en fonction de la représentation sur laquelle on insiste.
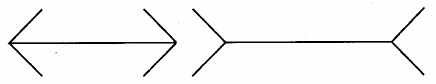 Figure de Müller-Lyer (les deux segments sont de même longueur)
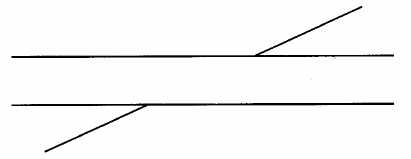 Illusion de Poggendorf
(une droite passe par les deux parallèles)
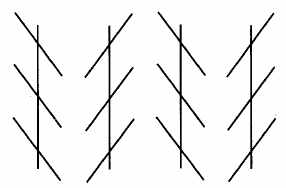 Illusion de Zollner (les
perpendiculaires sont parallèles).
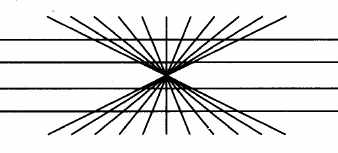 Figure en étoile de
Hering (les parallèles sont droites)
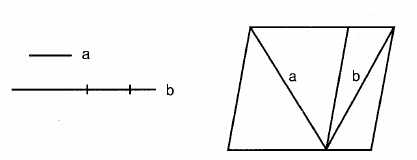 Illusion de Sander
Une surface grise apparaît plus claire à côté d’une surface noire qu’à côté d’une surface sombre. Il en va de même pour les couleurs. L’uniformité fondamentale du champ visuel, par exemple, se manifeste dans la tache aveugle et dans les déficiences du champ visuel, qui sont toujours complétées de telle manière qu’elles ne se remarquent pas dans la vision normale et ne sont généralement découvertes que par la périmétrie. Il s’avère que certaines figures correspondent à la dynamique figurative mentionnée. Fuchs (22) les appelle des figures contraignantes. Lorsque des points d’une figure circulaire apparaissent les uns après les autres et que l’un d’entre eux se trouve un peu en dehors de la figure circulaire, il est attiré dans celle-ci ; en cas de position excentrée, il se produit donc une distorsion en direction du centre. (Pour la « bonne » figure ou la « figure concise » de Wertheimer, voir Petermann (6)). En cas d’hémianopsie, une figure, par exemple un cercle ou une ellipse, est complétée lorsque la partie tombant dans la moitié saine du champ porte déjà en elle la loi du tout. Les figures compliquées ne sont pas vues malgré une représentation précise. Ce sont pourtant ces processus qui permettent la perception normale. Le noir, par exemple, n’est pas un stimulus sensoriel, mais une perception positive, rendue possible par la perception de l’environnement, par exemple lors de la lecture de cette écriture (23). La dynamique des proportions et des formes permet d’estimer normalement les objets. Lorsqu’il s’agit d’estimer les surfaces de rectangles, l’estimation devient plus précise si l’on se réfère aux proportions des côtés plutôt qu’à la comparaison directe des côtés individuels ; le plus précis est le nombre d’or. Les formes propres, par exemple comme le carré, se maintiennent face aux changements les plus forts, c’est-à-dire qu’elles ne peuvent être estimées que de manière imprécise ; en revanche, si les diagonales sont tracées, la sensibilité augmente immédiatement, les rapports de tension (voir l’illusion de Poggendorf) sont alors également vécus très fortement en termes de sentiments (25). L’organisation spatiale est également étroitement liée à l’organisation temporelle. A. Derwort a étudié les rapports de mouvement, la même chose étant valable pour les mouvements vus et exécutés. Un mouvement circulaire semble plus grand lorsque la vitesse augmente ; pour les figures, les courbes sont parcourues d’autant plus rapidement qu’elles sont étroites, c’est-à-dire qu’un cercle est déformé en conséquence lorsque la vitesse n’est pas régulière. Une figure est exécutée de manière inégale en fonction des courbes pour une vitesse subjectivement égale - vue ou exécutée. Si la courbe n’est pas vue, mais seulement suivie avec une petite lampe sur le doigt, alors la vitesse peut être uniforme. Si une ligne droite doit être décrite avec une vitesse croissante et décroissante, alors la ligne devient ondulante ; si la ligne droite est forcée, les mouvements sont hachés, c’est-à-dire que la forme temporelle est détruite (26). Le déroulement du temps lors de la perception et du mouvement est également structuré, c’est-à-dire que le futur et le passé sont inclus dans le présent ; dans le mouvement, la forme du temps se forme dans la figure. Alors que le déroulement physique du temps peut être géré de manière discontinue, le déroulement psychologique a la possibilité, lors de sa conception, d’englober en tout point des déroulements rétrospectifs et anticipés. Le champ de perception a une tendance à la structuration et à l’organisation. Plusieurs lignes parallèles, ce que l’on appelle la figure de Schumann (27), se présentent immédiatement comme des bandes ou des lattes avec un arrière-plan. Des séries sonores uniformes sont rythmées de manière subjective, le rythme pouvant être changé de manière totalement arbitraire (28). On les associe particulièrement facilement aux rythmes du pouls et de la respiration pour former un complexe particulièrement affectif. Lors de l’examen tachistoscopique, les complexes rythmiques peuvent être saisis beaucoup plus rapidement. Le fait que les processus de la Gestalt/forme soient causés par des processus cérébraux physiques, par exemple par des phénomènes électriques, comme l’avance Köhler comme théorie de la Gestalt/forme, s’avère déjà impossible sur des bébés aveugles opérés, comme le montre von Senden (9), et a également été réfuté dans ses points de départ par Petermann (6). Si l’on examine la formation des figures par tachistoscopie ou sur de petites figures, on constate qu’elles ne se forment pas de manière associative à partir des éléments individuels, mais qu’elles représentent un effort de différenciation à partir de figures organiques mobiles. Le premier élément est toujours la périphérie ou le contour, ce n’est que progressivement que l’espace intérieur se forme en direction du centre. On appelle ce stade la préfiguration. Wohlfahrt (29) a constaté sur de petites figures devenant lentement plus grandes qu’un cercle se forme d’abord comme contour ; l’intérieur est décrit comme une vie vacillante. Lorsque ce cercle s’étend sur des figures à l’apparence organique, qui sont en perpétuel mouvement, s’est consolidé jusqu’à la forme définitive (il est même décrit comme une enveloppe embryonnaire), c’est alors seulement que l’intérieur totalement indifférencié commence à se plastifier de manière élastique en noyaux et barrettes qui se dégonflent et se gonflent. Ce n’est qu’avec la structuration que cesse tout tissage et toute pression concentrés et agités, et que disparaît l’émotion forte provoquée par le fait de prendre forme. En tachistoscopie, c’est-à-dire en réduisant le temps d’observation à de très courtes périodes, le même phénomène se produit (30). Subjectivement, l’apparition de la forme a d’abord un caractère tout à fait affectif, très émotionnel (31), qui ne disparaît que lorsque la forme finale solide s’est formée. Conrad décrit cela comme suit : avec la formation complète, le « moi » se distancie de l’objet et devient libre grâce à cette distanciation. Il trouve la même chose dans la recherche perturbée de mots ; par exemple, les mots que l’on ressent de manière sourde (pour ainsi dire sur la langue) sont affectifs du point de vue émotionnel jusqu’à ce qu’ils sortent de la conscience de veille. Dans le processus pathologique, dans lequel le « moi » n’est pas libre face à ses contenus (idées fixes, etc.), les contenus sont restés au stade de la préforme. Dans le processus individuel, il apparaît donc que dans la représentation, la forme fixe se développe d’abord à partir de la forme mobile, puis de la partie individuelle. Plus la partie est considérée comme forte, plus le tout se désagrège. Cela correspond également aux processus organiques, qui présentent également un développement de forme mobile et fluide, à partir duquel les parties individuelles se différencient. Lors de la régénération de membres perdus chez les animaux inférieurs (par exemple les tritons), c’est la partie la plus périphérique qui se forme et se différencie en premier ; les parties plus centrales ne suivent qu’ensuite. Lorsque les formes se fixent, le processus de vie proprement dit régresse et, lorsque les parties deviennent indépendantes, l’organisme se dissout ou meurt. Aussi bien dans le développement des règnes animaux que dans celui de l’humanité et de l’enfant, on constate que la perception des parties individuelles ne se différencie que progressivement à partir de l’expérience d’ensembles ou d’images mythologiques (32). Sur des enfants, A. Heiss (33) expérimenta la capacité d’abstraction. Ils apprennent peu à peu à isoler des parties de complexes diversement structurés, et les illusions d’optique se montrent à un degré beaucoup plus élevé que chez l’adulte. Ce processus ne s’achève qu’à l’âge de 14 ans, date à laquelle commence réellement la capacité d’abstraction correspondant à celle de l’adulte. Köhler (34) n’a pu que très difficilement amener des chimpanzés à voir par exemple une branche sèche d’un arbre dans un but précis. Volkelt (35) a également constaté chez les araignées qu’elles vivent principalement dans des formes holistiques. Pour l’évolution de l’humanité, la naissance de la conscience philosophique à partir des représentations mythologiques est bien connue. Sander décrit maintenant des hommes à la vision plus globale et à l’observation plus analytique ; mais une nouvelle capacité est celle de voir à nouveau la partie en relation avec le tout, sans la détruire. Cette capacité ne peut pas être déduite de l’autre, car l’assemblage de parties ne donne que des constructions, pas de nouvelles formes. Si ce processus indispensable dans la vie normale est perturbé, par exemple par une lésion cérébrale, appelée protopathie par Conrad (36), le mot ne peut pas être trouvé à partir de l’épellation ; et si le mot est trouvé, il ne peut pas non plus être analysé en ses différentes lettres. La naissance de la forme s’accompagne d’un changement de conscience. La conscience claire de l’éveil et de l’objet, dont l’homme est libre d’affronter le contenu, n’a en fait que la forme finale aux contours fixes, en particulier lorsqu’elle est une partie isolée de la figure par opposition au fond. La forme préliminaire, dans la mesure où elle se présente déjà sous forme d’image, est vécue très fortement dans le sentiment et de manière presque onirique. Cela se différencie bien sûr dans les différents cas ; le fond n’est certes pas vécu de manière émotionnelle, mais pas de manière aussi consciente. Dans la mesure où le mouvement apparaît dans la perception, il acquiert un caractère plus émotionnel, de même l’imagination artistique a plus l’état de préfiguration onirique. La dynamique des forces dans les figures n’apparaît en fait que jusqu’au sentiment, comme on le constate régulièrement ; elle ne devient consciente qu’avec la représentation picturale. Le mode de pensée de Goethe constitue un prolongement particulier. Goethe décrit son idée de la plante originelle, tirée d’observations individuelles précises et variées, comme une représentation mobile, se métamorphosant constamment, à partir de laquelle il peut à tout moment développer des plantes possibles et réelles. Comme il l’a développée par le biais de l’observation partielle dans la pensée pleinement consciente, il s’agit d’une préforme d’un niveau supérieur, mais qui est activement acquise à partir de la forme finale fixe. En elle, la partie et le tout sont à nouveau réunis en une unité. La dynamique des processus de représentation décrite jusqu’ici représente certes les éléments de l’activité perceptive et montre dans quelle mesure des distorsions et des illusions naissent de la mise en valeur des forces propres ; mais cela ne suffit pas encore à expliquer l’acte perceptif proprement dit (37). L’homme n’a pas seulement des représentations sur la base d’impressions sensorielles, mais il est en mesure d’observer son environnement, il dépasse continuellement les limites de sa corporéité dans la perception (38). Pour la représentation, le propre corps est un objet comme les objets situés en dehors de lui, ils sont mis en relation mutuelle dans le champ de perception. Celui qui passe lentement à côté d’un autre train dans un train ne peut pas toujours déterminer avec certitude quel train est en train de rouler ; c’est sa propre liberté qui détermine quelle possibilité de représentation est réalisée, jusqu’à ce que d’autres points fixes en dehors des trains apparaissent dans le champ de perception ou qu’un changement de mouvement soit vécu par son propre organisme. Si, après un essai de rotation, par exemple sur une chaise pivotante, l’arrêt soudain se produit, il est connu que le mouvement de rotation en sens inverse s’installe avec nystagmus, etc. Ce phénomène est généralement rapporté aux processus mécaniques du système du canal semi-circulaire, les succès des mouvements ou les modifications du tonus sont considérés comme des réflexes de type typique. Mais si la rotation s’effectue par exemple dans un cylindre et que le sujet inverse soudainement la représentation, c’est-à-dire qu’il s’imagine au repos et que l’environnement défile, alors les mêmes phénomènes de réflexe ou de nystagmus se produisent, bien que seul le fait psychologique ait changé, et non les processus du canal semi-circulaire (39). Si le cylindre est déplacé en même temps que la chaise tournante, puis s’arrête, il n’y a aucune réaction, par contre tous les signes typiques de rotation apparaissent si seul le cylindre tourne et que le sujet s’adapte à la rotation propre. Les études de Christian (40) démontrent que l’impression sensorielle des organes vestibulaires n’agit vraiment que sur le champ de perception. Lorsqu’un sujet assis sur une chaise tournante dans l’obscurité est tourné imperceptiblement, c’est-à-dire de manière subliminale, que sa tête est fixée et qu’un point lumineux solidaire de la chaise tournante se trouve devant lui, le point lumineux commence à se déplacer, il est vécu comme un « point mobile sans changement de lieu ». Il indique avec précision la direction et le rythme du mouvement réel. Le scintillement autocinétique s’arrête immédiatement au début du mouvement. Lorsqu’un bourdonnement est perceptible, le sujet et le point lumineux semblent immobiles et le bourdonnement semble tourner, correspondant précisément au mouvement propre réel. Les images rémanentes se déplacent également en conséquence. Les images rémanentes sont également très sensibles à la stimulation rotatoire (41). Si, lors de la rotation, la relation entre la rotation propre et l’environnement — ce que von Weizsäcker appelle la cohérence — ne peut plus être maintenue, le vertige apparaît. Il est clair que la cohésion, c’est-à-dire la liaison imaginaire, est en cause ; le sujet ne peut plus inverser les mouvements, c’est-à-dire qu’il ne peut plus arrêter sa propre roue et laisser passer l’environnement. Le même phénomène se produit également pour le vertige optocinétique, c’est-à-dire lorsque ce n’est pas le sujet qui tourne, mais le cylindre qui l’entoure (42). Les malades dont la perception du corps est détériorée, les ataxiques, sont particulièrement vulnérables au vertige. - Les phénomènes de vertige cessent naturellement lorsque l’environnement tourne si vite que tout devient flou. Le vertige est un phénomène de cohérence tout à fait correspondant. En ce qui concerne la perception de l’équilibre, les sensations ne se transmettent pas directement aux organes de la réussite par la médiation des voies vestibulaires bien connues, mais elles ne sont transmises qu’au champ de perception ; dans la mesure où la posture et le mouvement s’orientent sur le champ de perception, les réflexes dits vestibulaires se manifestent. Ces processus dépendent de l’activité de représentation de l’individu. Ce n’est que grâce à cette activité qu’il est possible, en se déplaçant dans l’espace, de ne pas percevoir le mouvement des objets, mais de se déplacer soi-même. Cette faculté est également présente de manière excellente dans le règne animal et a été démontrée jusqu’aux abeilles et aux mouches (43). Si l’on a montré tout à l’heure que l’activité ou le mouvement est déjà nécessaire à la sensibilité tactile et à la représentation de l’espace en général, cette activité doit également être nécessaire pour que la perception normale des objets soit possible. C’est ce qu’exprime von Weizsäcker avec son « concept de performance ». Celui-ci est psychologique et inclut l’activité, sinon il s’agirait d’un concept technique qui signifierait le contraire de ce qu’il veut dire. Dans le « Gestaltkreis », von Weizsäcker explique plus en détail que la fluctuation du gonflement des stimuli et toutes les modifications de l’activité sensorielle provoquent certes des illusions et des déformations en ce qui concerne la représentation, mais que cette illusion permet la perception exacte et précise des objets. Il constate ce phénomène étrange que l’objet, dans la mesure où il est saisi, détermine l’irritabilité et les seuils des organes sensoriels de telle sorte qu’il soit perçu correctement. Si l’on appelle volonté l’activité psychique qui doit être saisie arbitrairement, il en résulte que la volonté s’empare de l’organisme physique et que l’activité des sens et de l’imagination fait plastifier les objets (44). Ce phénomène, selon lequel un homme qui se tient à deux mètres devant moi n’apparaît pas plus grand que quelqu’un qui marche à trente mètres de distance, selon lequel les objets apparaissent constants malgré les changements de distance et d’éclairage, est appelé la « constance des choses vues », très étudiée. Elle se rapporte aussi bien à la perception spatiale qu’à la perception visuelle. Une surface grise en pleine lumière, qui émet objectivement plus de lumière qu’une surface blanche à l’ombre, apparaît néanmoins grise (46). Si l’on regarde par exemple, d’un seul œil, des bâtons qui nous font face en biais, de sorte qu’ils sont réduits par la perspective mathématique, ils obtiennent dans l’évaluation leur longueur normale, dans la mesure où l’environnement peut également être perçu ; s’ils apparaissent par contre comme des traits lumineux dans l’obscurité totale, l’observateur ne peut que s’en tenir à la perspective mathématique et en arrive à une erreur d’appréciation objective (47). Les rails se rapprochent les uns des autres dans la perspective, mais on remarque tout de suite si leur distance objective augmente en s’éloignant. Cela n’est pas seulement dû à la séparation transversale des parties de la rétine, car celles-ci n’ont pas de correspondance fixe : d’un point de vue purement physiologique, il devrait en résulter un enchevêtrement chaotique, sans compter que les valeurs initiales peuvent se déplacer dans des cas pathologiques (perte partielle du champ visuel). Von Weizsäcker a étudié ces rapports avec précision. Ce phénomène est particulièrement bien mis en évidence par les sensations de pression. Pour des poids particulièrement importants, on peut constater certains seuils exacts et constater la dépendance de l’estimation du poids par rapport aux moments mécaniques. M. von Frey a étudié les rapports entre les poids soulevés et l’estimation réciproque du bras de levier (48). Un tube a été placé sur le bras, après quoi le poids estimé correspondait exactement au bras de levier (49). Mais si l’on crée des conditions dans lesquelles le mouvement peut s’affirmer face à la mécanique, on obtient bien sûr des rapports totalement différents. Un poids est estimé de la même manière avec les dents, le pied, le cou et le bras, qu’il s’agisse de la main, de l’avant-bras ou du bras supérieur (50). Si l’on peut admettre que le mouvement s’oriente selon la perception, alors le poids est perçu comme la gravité par excellence, par opposition à la tendance à se redresser, car le champ de perception fonctionne comme un tout. Or, la perception peut changer immédiatement : on peut ressentir le poids du bras même si celui-ci n’est pas chargé, alors qu’il est éliminé par ailleurs, et en tenant un objet, l’attention peut aussi se concentrer sur la pression exercée sur la peau. Lorsqu’un poids de 2 kg est porté, il en résulte la même estimation que lorsqu’il tire sur une poulie sur le bras couché (51). Mais un deuxième objet peut également être éliminé sans problème ; si l’on place un poids dans un panier, il ne semble pas plus lourd que si on le prend ensuite à main nue. La tare est également impliquée en permanence dans les mouvements, par exemple dans le jeu de tennis. C’est l’illusion permanente de l’état intramusculaire qui permet de se prononcer correctement sur l’objet (52). Lorsqu’une personne atteinte de la maladie de Parkinson présente une rigidité musculaire, le patient doit subjectivement faire des efforts de volonté beaucoup plus importants que la normale, par exemple pour bouger son bras. Si l’on tient le bras d’une telle personne, il semble que la force nécessaire pour le fléchir soit beaucoup plus importante que chez les personnes normales, alors qu’en réalité, c’est l’inverse, comme on peut le constater par des mesures. On perçoit donc directement l’effort de volonté de l’autre (53). On peut donc effectivement avoir accès à l’âme d’un autre être humain par la voie des sens, grâce à l’empathie ! Le fait que cela doive d’abord être interprété à partir d’indices extérieurs est une théorie erronée ou un cas particulier ! Christian et Pax (54) ont pu démontrer, à l’aide de processus oscillatoires, que les oscillations pendulaires sont vécues par exemple comme un mouvement régulier qui s’arrête brièvement à chaque point de retour. L’impulsion de mouvement, dont la confrontation avec la force de gravité est à l’origine de l’oscillation pendulaire, est extraite du processus global. C’est ainsi que presque toutes les oscillations sinusoïdales sont vécues dans leur projection mathématique sur la ligne droite, c’est-à-dire comme des impulsions de mouvement entre des polarités dynamiques. Dans le cas du goût, par exemple, les sensations de goût, d’odeur, de température et de toucher des parties molles et des dents interagissent. Lorsque l’on entaille du bois avec un scalpel pointu sous une très forte loupe ou un microscope, il nous apparaît doux comme de la cire, parce que la perception visuelle est déformée par les conditions optiques et n’est pas réduite dans l’imagination (55). Dans le processus de perception pleinement conscient, nous n’avons qu’une capacité de compréhension limitée. Dans le champ visuel, seule une petite partie autour du point de fixation peut être saisie de cette manière. Plus nous nous tournons vers l’observation analytique partielle, plus ce champ devient étroit et peut se réduire à un point. Cet élément ponctuel, qui peut apparaître dans tous les domaines sensoriels et dont la psychologie atomiste pensait qu’il composait tous les complexes supérieurs, est un résultat final d’un effort de représentation. Il n’est possible de saisir que jusqu’à 5-6 parties ou points individuels dans un acte de perception, peu importe qu’ils se trouvent l’un à côté de l’autre ou qu’ils ne puissent être vus que par un mouvement des yeux (56). L’atome psychologique est un produit, pas une donnée. Si des groupes d’objets sont ordonnés rythmiquement (57), on peut en voir davantage, par exemple 3 x 3 ou 4 x 3, mais il s’agit alors de deux activités l’une dans l’autre. Parmi les sommets de figures géométriques régulières situées dans le cercle, seules 7 formes peuvent être appréhendées, le triangle régulier jusqu’à l’octogone et le cercle, les autres étant des polygones réguliers (58). Ce champ de perception s’élargit à mesure que l’on envisage des figures ou des relations globales. Mais l’étroitesse de ce champ est entièrement déterminée par l’objet et non par l’angle visuel, c’est-à-dire que l’angle visuel pour ce champ diminue avec la distance. Ce rétrécissement concentrique du champ visuel est appelé le phénomène d’Aubert-Förster (56). L’activité sensorielle de l’homme ne peut en aucun cas être présentée comme une simple continuation de la stimulation, si l’on ne veut pas se mettre en contradiction avec les faits ; la théorie qui veut expliquer, à partir de la connexion interneuronale (60), la distorsion et la dépendance spatiales et temporelles des perceptions et des mouvements sur la base des résultats expérimentaux lors de la stimulation artificielle des nerfs, peut considérer la perception réelle comme statistiquement possible — dans la mesure où il y a des illusions, etc. — mais ne peut jamais justifier sa constitution. Jusqu’à présent, on a donc trouvé : 1. l’activité sensorielle est imprégnée idéellement, sans cette activité de représentation, il n’y a que des sensations, pas de perceptions. Ce seul fait rend impossible une explication mécaniste ou atomiste. Pavlov (61) ne tient pas compte, dans la justification des réflexes conditionnels, du fait que les stimuli déclencheurs contiennent déjà la plupart du temps des relations idéelles. Même sa doctrine sur l’entraînement et l’inhibition apparaît sous un autre jour si l’on tient compte de ce fait. Si, par exemple, il produit un réflexe à un endroit précis suite à un stimulus de contact, mais qu’il ne parle pas de la sensation, mais de la localisation, et qu’il stimule ensuite d’autres points après lesquels la nourriture n’apparaît pas, par exemple, le réflexe s’éteint tout d’abord complètement parce que le chien est irrité ; c’est ce qu’on appelle alors l’inhibition. Ce n’est qu’après une pratique fréquente que l’animal apprend à réagir à l’endroit localisé, mais il s’agit là d’un effort de représentation. Si les réflexes conditionnés ne sont plus possibles en raison de l’ablation des centres cérébraux secondaires nécessaires à la pénétration idéelle du champ sensoriel, ce phénomène ne doit donc pas être interprété uniquement dans le sens des théories de la conduction. 2. Pour l’espace qui apparaît dans le champ de perception, la continuité doit être établie, contrairement à l’espace discontinu et illimité de la grille d’impulsions de la physique (62). Il faut distinguer l’espace tactile, l’espace actif et l’espace visuel (63) ; ces espaces sont également construits de manière légèrement différente. Pour l’espace visuel, il y a d’abord dans l’obscurité une surface à deux dimensions, qui est déplacée dans l’acte visuel proprement dit avec l’apparition de la troisième dimension, mais au fond, elle est toujours présente comme limite de l’espace. D’un point de vue géométrique, cet espace se présente comme non euclidien (64) et correspond le mieux aux représentations spatiales de la géométrie synthétique (géométrie projective ou géométrie de position) (65). Contrairement à la géométrie euclidienne, la géométrie synthétique est une géométrie non métrique, en fait purement qualitative, à partir de laquelle on peut cependant développer la géométrie analytique, métrique. Staudt n’a eu besoin d’aucune notion de base de la géométrie élémentaire pour sa « géométrie de la position ». Cet espace est certes infini, mais pas illimité ; dans la périphérie infinie, l’espace devient surface, la surface devient ligne, la ligne devient point. L’espace visuel peut être considéré comme un tel espace, limité par le fond comme par la périphérie infinie. Cette géométrie s’applique aussi parfaitement à la conception organique, comme l’ont montré à plusieurs reprises George Adams et L. Locher-Ernst. Le temps n’est pas non plus le temps discontinu de la physique. Il peut tout d’abord s’écouler à des vitesses très différentes, c’est pourquoi on a également parlé d’un temps biologique (66). Dans le présent, le passé et le futur sont fonctionnellement co-créateurs. L’espace et le temps ne se comportent pas simplement comme un système à 4 coordonnées, mais comme une structure cohérente dans laquelle ils peuvent se représenter mutuellement dans une certaine relation (67). Lors du toucher d’objets, par exemple, le mouvement qui se déroule dans le temps conduit à une représentation spatiale. 3) Si l’on établit les conditions des illusions perceptives, il s’avère qu’une dynamique de forces est inhérente aux configurations. Si cette dynamique ne peut pas toujours s’exprimer de manière imagée lors de l’observation ordinaire, elle permet néanmoins dans une large mesure, comme cela a été démontré lors de l’estimation de rectangles, d’observer et de juger avec précision, même si elle ne constitue pas la perception finale. 4) Si l’on examine la formation des représentations ou des formes spatiales dans des conditions appropriées, la théorie de la construction additive se révèle inexacte. Ce sont toujours les formes entières et uniformes qui sont primaires et à partir desquelles les parties sont différenciées. La forme préliminaire a d’abord un caractère plastique, elle est en perpétuel mouvement, elle se dessine rapidement de la périphérie vers le centre, ce n’est qu’après avoir été façonné qu’elle devient fixe dans la forme finale. Elle porte entièrement le caractère de la création organique. Ce n’est qu’à partir du point de fixation et de la pièce détachée qui en résultent que l’on peut construire des complexes dans le sens de la technique et de la mécanique. Lorsque des points se déplacent sur un fond indifférent et que la forme est arbitraire pour l’imagination, des relations se forment dans le sens des lois physiques, par exemple selon les lois de Kepler. Si la forme ne peut pas devenir fixe en elle-même, la dynamique donne naissance à la mécanique (68). En cas de lésion du système nerveux, ce ne sont pas des éléments individuels ou des relations complexes qui font défaut, mais la capacité de différenciation est perturbée. La spécification diminue et le collectivisme augmente (69). 5. Avec la formation de la forme, le rapport psychologique change également : la préforme agit le plus fortement de manière affective, c’est-à-dire sur la volonté, au début de la formation imagée ou juste avant ; la plasticité en mouvement, le mouvement et la dynamique en général vivent principalement dans le sentiment, et seule la forme finale, solide, est objectivée et devient une véritable « représentation ». L’homme se trouve alors face à elle. En même temps, l’état de conscience s’accroît. Ce n’est que la forme finale détaillée qui est intégrée dans la pleine conscience de veille. Comme nous l’avons montré avec Goethe, la représentation peut être déplacée par l’activité et acquérir le caractère de préforme, mais de manière exacte à un niveau plus élevé que la pleine conscience de veille. 6) L’activité est nécessaire de différentes manières pour que cela se produise. L’activité de la volonté se manifeste soit seulement dans la représentation, dans la « plastification » des objets, soit directement dans le mouvement. Elle envahit en fait constamment l’organisme et, selon l’objet avec lequel elle entre en cohérence, les seuils des organes sensoriels et l’attribution des points sensoriels sont modifiés. L’orientation par rapport à l’environnement en cas de mouvement propre n’est possible que grâce à ce fait psychologique. Ainsi, les processus de l’environnement, y compris les processus psychiques des autres personnes, peuvent être vécus directement, si la disposition et la possibilité pour les représentations correspondantes sont présentes. von Weizsäcker désigne cette imbrication de la perception avec l’activité motrice et la volonté par l’image symbolique du « cercle de gestalt ». Le principe de performance postulé par Weizsäcker ne doit pas être pensé de telle sorte que les objets dans l’espace agissent directement sur l’homme, car c’est sa propre activité qui s’empare des objets. Cette saisie est donc une énigme, même si elle existe en tant que fait. Le même problème, mais à une échelle beaucoup plus grande, est celui de l’orientation dans l’espace sur de plus grandes distances. C’est surtout le cas des oiseaux migrateurs, mais d’autres animaux et même, dans certaines conditions, l’homme, peuvent s’orienter vers des endroits précis qu’ils ne perçoivent pas du tout. Les abeilles peuvent être dressées à des endroits ponctuels très précis dans l’espace, même s’il n’y a aucune possibilité d’orientation à des kilomètres à la ronde. Les résultats sont si étranges que l’espace pourrait être pensé de manière qualitativement structurée, de sorte que les signes locaux de Lotz ne paraissent pas si absurdes. Jusqu’à présent, ces phénomènes ont résisté à toutes les tentatives d’interprétation physique (70). – Pour les animaux, la dépendance de la perception à l’activité motrice joue un rôle particulièrement important. Buytendijk (71) rapporte qu’un chien apprend plus facilement à distinguer des figures lorsqu’il peut se déplacer sur celles-ci. Lashley (72) a exercé des rats sur des lignes verticales et horizontales au cours de 500 à 800 expériences, sans parvenir à une preuve de la capacité de distinction, qui se manifestait immédiatement lorsque c’était des objets vers lesquels ils devaient sauter. Du côté des sens, la prémisse du « parallélisme psychophysique » selon laquelle les organes de la réussite pourraient être excités directement par les organes des sens via les voies de conduction connues ne s’applique pas. La relation stimulus-réaction prend son chemin à travers le psychisme lui-même. Mouvement Après avoir examiné le côté sensoriel, il faut examiner si le mouvement doit être compris comme une mécanique — plus ou moins compliquée. Le mouvement peut tout à fait être réglé par des centres et l’action des membres peut être comprise à partir de la mécanique du système ostéo-musculaire. Subjectivement, le mouvement a cependant la primauté. Du point de vue de la perception, le mouvement et la mécanique se distinguent tout à fait.Ils se distinguent fondamentalement par le fait que le mouvement repose sur une activité créatrice. Si l’on agit dans un contexte purement idéel, on a pour lui une impulsion immatérielle clairement compréhensible. L’organisme qui se meut spontanément (73) est en contradiction avec le principe d’inertie qui s’applique à la matière inanimée. Si l’on établit la condition de grandes accélérations, on peut démontrer que l’homme est également soumis à l’inertie. Mais cela ne vaut que pour son organisation matérielle ; grâce à l’activité psychique, le corps est arraché à l’inertie, mais un homme devient psychiquement inerte lorsqu’il s’abandonne passivement à l’organisation matérielle. En y regardant de plus près, on constate également que le mouvement se maintient certes dans les limites de ce qui est mécaniquement possible, mais qu’il est soumis aux conditions d’espace et de temps telles qu’elles sont décrites pour le psychologique. La possibilité que les mouvements soient dirigés par des centres du système nerveux central a été abondamment étudiée par Bethe dans ses études sur la plasticité du système nerveux (74). Les mouvements d’un organisme sont tout d’abord tous coordonnés entre eux (coordination). Cette coordination se réfère maintenant tout à fait à l’environnement, comme dans la course, la marche, etc. L’homme apprend d’abord ses capacités, alors que chez l’animal, la coordination complète apparaît très rapidement. Différents auteurs se sont demandé si la coordination des mouvements entre eux et leur relation à l’objet devaient d’abord être apprises par le système nerveux ou si la coordination faisait partie de la nature des mouvements et était présente dès la capacité de mouvement. 153 Coghill et Herrick (75) ont pu montrer que chez l’embryon, il existe déjà des mouvements spontanés complets, sans qu’ils puissent être interprétés comme une réactivité. Selon des études histologiques, il n’existe pas encore de connexions entre les neurones sensoriels et les neurones moteurs à un stade du développement où de telles activités sont déjà présentes. Weiss (76) décrit que lors de l’éclosion des poules et des papillons, des mouvements ordonnés sont présents dès le départ. Il a ouvert des femelles salamandres dans l’utérus desquelles se trouvaient des embryons de 2 cm de long, serrés et plus ou moins refoulés, sans possibilité de mouvement. Lorsqu’on les jette dans l’eau, ils nagent tous immédiatement normalement et avec un jeu de mouvements coordonnés. Selon Graham Brown (77), lorsque des fœtus de chat à naître sont placés dans une solution saline physiologique chaude, on peut observer des mouvements de cris bien coordonnés et obtenir des réflexes clairement coordonnés en réponse à des stimuli. Carmichael (78) a laissé des germes d’axolotl se développer dans une solution de chloroforme et d’acétone. Le développement morphologique, comparé à celui d’animaux témoins dans de l’eau normale, s’est déroulé normalement. Le système nerveux était donc sous anesthésie permanente et aucun mouvement n’était visible. Après un transfert ultérieur dans l’eau du robinet, les animaux ont mis environ une demi-heure à se réveiller, puis ils se sont mis à bouger avec autant d’assurance et d’agilité que les animaux de contrôle qui avaient pu expérimenter les différents stades de mobilité. Des expériences analogues ont été réalisées par Matthews et Detwiler (79) (larves d’amblystes dans du chloréthane). L’apparition de la capacité de mouvement s’accompagne d’une formation homogène et cohérente, c’est-à-dire d’une relation structurée avec l’environnement. Il est cependant encore possible que les actions soient préformées dans des mécanismes centraux. Tout d’abord, des réponses précises à des stimuli peuvent être données avec des membres inhabituels, si les membres correspondants sont empêchés ou occupés par une action plus importante. Szymanski (80) appelle ce comportement le « principe du déroulement non perturbé de l’action préférée ». Si l’on chatouille le nez d’un crapaud pendant le réflexe d’agrippement, il utilise sa patte au lieu de son bras habituel. Une grenouille décapitée qui réagit avec sa jambe à des stimuli sur la peau de l’abdomen par ce que l’on appelle le réflexe d’essuyage utilise immédiatement la jambe 154 du côté opposé lorsque l’autre est maintenue. Cela peut être observé en détail lorsque l’homme est dérangé dans des activités qui l’occupent beaucoup. Lorsqu’on attache un chien, il fait des mouvements de dégagement sensés, bien qu’il n’ait jamais été amené à faire de tels mouvements auparavant. Bâytendijk a examiné si les mouvements normaux et particuliers devaient être interprétés comme des réactions en chaîne (81). La conception des mouvements normaux en tant que réflexes en chaîne, défendue auparavant, a d’abord été clairement réfutée par Graham Brown (82) et plus tard par Holst. Brown a coupé les nerfs sensitifs de chiens spinaux et a tout de même obtenu des mouvements de marche. Holst a coupé les racines postérieures des poissons, mais la coordination des mouvements des nageoires n’en a pas été affectée (86). Les coléoptères aquatiques, qui normalement n’utilisent que leurs pattes arrière pour nager, exécutaient immédiatement les mouvements de nage corrects avec la patte suivante après l’ablation d’une patte arrière ; si l’ablation n’était que partielle, la seconde intervenait dans la mesure nécessaire. Les animaux avec un grand nombre de pattes, comme les mille-pattes, ont une coordination fixe et bien déterminée des pattes. A chaque nouvelle ablation de certaines pattes, de nouvelles combinaisons et coordinations, jamais réalisées auparavant, se mettent immédiatement en place. Si un centre déterminé devait exister pour chaque type de coordination, il s’avère déjà quantitativement, par le nombre de transformations possibles, que cela n’est pas du tout possible. Si l’on voulait s’en tenir à l’hypothèse des centres, il faudrait déjà un nombre excessivement élevé de centres pour les différents types de mouvements normaux. Bethe a trouvé 255 combinaisons possibles rien que chez les opilions (tisserands). Le passage de la cadence quadrupède à la cadence tripode chez le coléoptère aquatique Dytiscus marginalis a lieu même après la suppression du cerveau et des 1er et 2e ganglions pharyngiens. Buddenbrock (83) a fait des expériences similaires avec le criquet bâton Dixippus morosus. Dans le cas de l’étoile de mer, qui est couchée sur le dos et se retourne, le nombre mathématiquement possible de remaniements (Bethe) se réalise également après l’enlèvement de bras. L’étoile serpentine se déplace comme un flagellaire avec un seul bras après avoir perdu les quatre autres (84). Les chiens aussi adoptent des formes de locomotion très différentes avec la perte de leurs membres. On sait qu’ils peuvent à tout moment passer de la marche quadrupède à la marche tripode 155 . La perte de tous les membres entraîne un tout nouveau mouvement de reptation, semblable à celui des chenilles (Bethe). Plus on enlève de membres, plus le rythme des autres s’accélère, de même qu’ailleurs, plus la taille du corps diminue, plus le rythme des mouvements s’accélère (85). Si l’on raccourcit des poissons ou des vers, on retrouve le même phénomène (86). Si l’on morcelle des animaux inférieurs, comme les planaires (vers tourbillonnants) ou autres, chaque morceau donne naissance à un animal entier. Les animaux légèrement supérieurs, comme les amphibiens par exemple, remplacent les membres perdus. Ce phénomène est très marqué dans les stades embryonnaires antérieurs et a conduit à la notion de champ en biologie (pour plus de détails, voir Spemann (87)). Il en va de même au niveau fonctionnel : si l’on coupe en deux des vers (88), des serpents ou des anguilles, chaque moitié se déplace de manière autonome comme un animal normal de longueur correspondante, également en fonction du rythme et de la longueur du mouvement d’ondulation. Or, on connaît chez différents animaux ce que l’on appelle l’autotomie (si l’on tient par exemple un lézard par la queue, ce bout de queue se détache, si l’on saisit à nouveau le moignon, le suivant, etc.) Chez le ver de terre, l’autotomie progressive peut se faire jusqu’à l’extrémité de la tête, mais si on le divise auparavant, l’extrémité antérieure de la partie postérieure séparée forme fonctionnellement une tête, et le découpage progressif ne peut se faire que jusqu’à cette tête fonctionnelle, alors que cette limite n’existait pas chez l’animal non divisé (89). Mais ce phénomène ne dépend pas non plus du cerveau lui-même, car les lézards s’autotomisent même après décérébration. Thorner appelle cela une « réaction holoplastique » (90). Or, il y a une chose étrange : si l’on relie mécaniquement un ver de terre divisé à ses parties coupées, la coordination de l’animal entier se rétablit immédiatement (91). Cette constatation, qui peut être proche de certaines formations d’états dans le règne animal, comme par exemple chez les siphonophores (méduses d’état), peut également être constatée chez les animaux supérieurs après section de la moelle épinière. On sait en effet que la moelle épinière ne peut pas se régénérer, toute séparation est définitive. Naunyn, qui a étudié les questions de régénération (92), s’est laissé tromper chez les jeunes chiens en interprétant la restitution des mouvements après 4 à 5 jours comme une régénération ; Schieferdecker (93) a ensuite démontré qu’il s’agissait de réflexes coordonnés de l’animal postérieur, déclenchés par l’animal antérieur. Un grand nombre d’études ont confirmé que la coordination des mouvements peut se rétablir de manière tout à fait normale malgré la section de la moelle épinière : I. Steiner l’a étudié sur des animaux inférieurs et des poissons (94), ten Cate sur des requins (95), Bickel sur des anguilles (96) ! Hooker sur le poisson rouge (97), Babak sur de jeunes larves de grenouilles (98) ; vérification par Hooker (99), qui a également poursuivi les recherches sur d’autres jeunes animaux et animaux inférieurs (100) et a coupé la moelle épinière intra-utérine avec de jeunes embryons de rats de Nicholas (101) ; Bikkel sur la grenouille (102). Chez l’anguille, il a seulement constaté que la moitié de la moelle épinière coupée avait perdu la capacité de nager en arrière et ne pouvait pas non plus s’intégrer dans les mouvements correspondants de l’animal antérieur. Il a coupé la moelle épinière de tortues plus petites à différents niveaux, mais n’a pas obtenu de troubles moteurs (103). Spatz, qui voyait également chez de jeunes animaux l’activité réflexe du membre postérieur à travers des mouvements volontaires des extrémités antérieures, de sorte que des actes moteurs presque normaux apparaissaient (104), a en outre démontré que la moelle épinière sectionnée présentait également des réflexes normaux et nouveaux (105). Les résultats obtenus par Goltz et Freusberg (106) ont été vérifiés par Philippson (107), qui a enregistré les mouvements d’appui, de course et de galop par voie cinématographique. Bien qu’il ne s’agisse pas de réflexes en chaîne (82, 86), la coordination du mouvement ou le mouvement lui-même n’est rendu possible que par l’activité de perception. Bickel a coupé les racines postérieures d’une grenouille qui n’exécutait des mouvements coordonnés qu’après la section de la moelle épinière. Comme l’afférence sensitive était interrompue, l’animal postérieur était traîné mollement (96) ; en cas de section unilatérale, une moitié était paralysée, mais après quelques jours, la motilité se développait à nouveau progressivement dans la jambe sensitive. En outre, une grenouille dont la moelle épinière a été sectionnée se déplace mieux sur un sol rugueux que sur un sol lisse. La même constatation a été faite par Snyder (108), dont les salamandres furtives semblaient avoir une coordination des mouvements presque inchangée, alors qu’elles ne bougeaient pas sur une surface lisse. Quelle est la position fonctionnelle de la musculature dans le mouvement ? Comme le montre l’étude approfondie de Baeyer (109), les différents muscles peuvent remplir des fonctions très différentes en fonction des mouvements. Le même muscle peut agir comme fléchisseur et comme extenseur, deux muscles peuvent se comporter comme synergistes et antagonistes selon les conditions ; un seul muscle peut s’intégrer dans une action avec une fonction unique, mais aussi différencier ses différentes parties pour différentes tâches. Ainsi, même pour le mouvement normal, les différents organes sont intégrés avec des fonctions différentes. Si l’on déplace maintenant des muscles ou leurs tendons, ils sont immédiatement intégrés correctement dans leur nouvelle position. Perthes (110), en cas de paralysie du radius, a déplacé le flexor carpi radialis et ulnaris du côté de l’extension et a suturé le premier avec les tendons des trois extenseurs du pouce et le second avec l’extenseur digital commun. La fonction est revenue si bien que le patient a pu jouer du piano et du violon sans problème. Cette opération, qui n’est qu’un exemple, a été répétée à plusieurs reprises. De nombreuses greffes de tendons ont été effectuées de la même manière (111). Il est intéressant de constater que la prise en charge des nouvelles fonctions s’est faite très soudainement, de sorte qu’un lent réapprentissage des centres n’entre pas en ligne de compte. Ce phénomène est particulièrement bien illustré par les opérations des muscles oculaires. Monzardo (112) a raccourci et allongé les muscles oculaires d’animaux de laboratoire ; entre autres, l’un des muscles oculaires droits a été divisé et une pointe a été attachée à 90 degrés sur le côté. Bien que ces interventions, qui perturbent fortement la mécanique, ne puissent être que très difficilement compensées sur le plan fonctionnel, les modifications ont été corrigées au bout de quelques jours. Marina (113) a remplacé le muscle rectus internus par le muscle obliquus superior ou a interverti les deux recti — internus et externus ; le muscle rectus internus a été remplacé par le rectus superior. Lors de ces expériences sur des singes, lorsque le bandage était retiré au bout de 3 à 4 jours, la position correcte des yeux et le nystagmus rotatoire correct étaient immédiatement présents des deux côtés. Selon Jackson (114), le muscle oblique supérieur paralysé peut également être remplacé par le déplacement de l’insertion du muscle droit supérieur, de même que le muscle droit externe paralysé par la séparation du muscle droit supérieur et du muscle droit inférieur avec ou sans ténotomie du muscle droit interne. Après une amputation, on sait que la formation de tubes musculaires entraîne également un changement de fonction de la musculature, qui s’effectue sans problème (voir Bethe (74)). 158 Pour le mouvement, il n’est pas possible de penser qu’un centre travaillant automatiquement excite les différents muscles et que l’image du mouvement qui apparaît représente le résultat plus ou moins fortuit ou la résultante des contractions musculaires. Au contraire, chaque muscle, quelle que soit sa position aléatoire, est intégré dans la forme du mouvement, à moins d’une impossibilité mécanique. Le mouvement lui-même s’oriente vers les organes donnés ou se plastifie à partir de là. La coordination des membres dépend du nombre total de membres présents, la forme du mouvement dépend des membres présents. Cela est particulièrement visible chez les étoiles-serpents, mais aussi chez les chiens, où l’amputation permet d’obtenir des formes de mouvement entièrement nouvelles, par exemple le saut de kangourou (115). Lors de la division d’animaux, deux animaux entièrement nouveaux sont créés ; cela se produit immédiatement sur le plan fonctionnel — si l’animal est encore viable — et, chez les animaux inférieurs, également sur le plan morphologique par régénération. Lorsque les moitiés divisées sont reliées, les mouvements s’organisent à nouveau en un tout homogène ; cela se produit chez les animaux supérieurs après la section de la moelle épinière. L’une des parties doit cependant pouvoir percevoir les mouvements de l’autre, bien qu’en cas de moelle épinière intacte, le mouvement soit possible même après resensibilisation. Pour la perception, il a été démontré que la performance, la saisie de l’objet d’une part et la conception d’autre part doivent être différenciées. Pour le mouvement, il doit donc aussi paraître légitime d’examiner si la performance n’est possible qu’en raison de la conception, c’est-à-dire si elle doit être différenciée de celle-ci. Dans les études de Derwort sur les mouvements figurés, ceux-ci présentent la même régularité que les figures perçues (116). Les mouvements conjoints sont un phénomène bien connu. Le principe de performance ne peut pas leur être appliqué, car ils sont superflus pour la performance purement objective. Bethe a alors étudié le rapport entre ces mouvements conjoints et la dépendance des membres les uns par rapport aux autres. Le coléoptère aquatique Dytiscus est clairement inhibé sur terre par la fixation de l’articulation de la hanche, dans l’eau il est totalement impuissant, ce n’est qu’en cas d’amputation que la patte centrale entre en action pour nager. Si les cuisses des pattes sont collées au corps de l’insecte, celui-ci ne peut pas se déplacer, mais 159 seulement se débattre par crises pendant des jours (117). Chez le chien, le fait d’attacher une jambe ne suffit pas encore, car il est encore possible de faire des mouvements. Mais dès qu’une patte est plâtrée, les troubles moteurs les plus graves apparaissent ; lorsque l’animal est au sol, il ne fait que se débattre et ne peut pas se redresser. S’il ne reste qu’un moignon de jambe après l’amputation, celui-ci exécute également les mouvements conjoints, le plâtrage entraînant ici aussi de graves troubles. Si les racines postérieures d’une jambe plâtrée sont coupées, les troubles diminuent certes, mais une gêne subsiste, qui cesse immédiatement après la libération. En revanche, en cas de section de la racine antérieure, c’est-à-dire d’interruption des voies motrices, l’immobilisation n’entraîne aucun handicap, au contraire, l’animal n’est que peu gêné par la jambe qui traîne mollement (118). Chez l’homme, l’immobilisation d’un membre entraîne également une gêne plus importante que celle qui se justifie d’un point de vue purement mécanique ; même un bras plâtré gêne la marche. De même, une performance physique intense, par exemple une course rapide, nécessite un mouvement non entravé. Le mouvement a tout d’abord un caractère de champ, ses différentes articulations sont toujours en relation les unes avec les autres ; les rapports organiques d’espace et de temps s’appliquent à celles-ci. La relation de l’organisation du mouvement est une relation motrice qui est renforcée par la sensibilité, mais qui n’est pas constituée. Si la sensibilité est entièrement conservée et que seule la motricité est interrompue, le membre sort du contexte du mouvement. La performance dépend alors de l’organisation non perturbée du mouvement. Selon les explications de von Weizsäcker, le fait psychologique de la cohérence est toujours nécessaire pour une performance, c’est-à-dire que les tendances propres du mouvement sont subordonnées à l’intention. Mais si l’on crée des conditions dans lesquelles elle s’exprime, il est possible de faire quelques constatations. Lorsqu’une personne pense aller tout droit dans l’obscurité ou sur un terrain sans possibilité d’orientation particulière, elle décrit un cercle, c’est bien connu. Ce fait est en effet attribué à l’intensité de mouvement un peu plus forte d’une jambe. D’un point de vue mécanique, cela est vrai, mais si l’on considère le résultat d’une longue marche dans les aléas physiologiques, il est impossible, ne serait-ce que du point de vue statistique, que le cas particulier, à savoir le cercle, se produise toujours. D’un autre côté, le cercle est une forme fondamentale de l’organique. Lorsque les animaux en captivité (119) se déplacent en raison de leur instinct d’activité, mais que les grilles les empêchent d’entrer dans une cohérence naturelle avec leur activité, des mouvements stéréotypés apparaissent, par exemple le tissage bien connu des ours, des éléphants, etc. Si l’on observe attentivement le tissage de différents animaux, on peut également y trouver la lemniscate comme forme de base extrêmement fréquente, même si elle est modifiée de différentes manières. Les mouvements circulaires peuvent être obtenus expérimentalement par des stimuli unilatéraux, par exemple des stimuli labyrinthiques ou des lésions cérébrales unilatérales, par exemple du cervelet. Dans certains cas, le champ de mouvement peut toutefois conserver la tendance lorsque la cause du stimulus a disparu. I. Steiner (94) a obtenu des mouvements forcés circulaires chez la roussette (Scyllium canicula) par ablation de la base du mésencéphale. Si l’animal a ensuite nagé en cercle pendant au moins 10 heures et qu’il est décapité, il poursuit le mouvement circulaire. En revanche, une lésion unilatérale de la moelle épinière ne permettait pas d’obtenir un mouvement circulaire. R. Dubois (120) a fait la même expérience un peu plus tôt avec des coléoptères (avec Dytiscus marginalis). Steiner a ensuite répété l’expérience sur les coléoptères avec le même résultat chez Carabus auratus. En enlevant la moitié du ganglion dorsal, le coléoptère a été contraint à un mouvement circulaire ; après deux minutes de course, le coléoptère a été décapité, mais l’animal a conservé son mouvement circulaire. Les expériences sur les requins ont été confirmées par Bethe (121). Mais il a également constaté que la position asymétrique des nageoires en cas d’extirpation unilatérale du labyrinthe persiste après la section de la moelle épinière et que l’effet positif peut se transformer en effet négatif (122). Il avait également observé des phénomènes après des extirpations labyrinthiques unilatérales et bilatérales, qui étaient en contradiction avec les conceptions classiques d’Ewald (123). Après l’ablation d’un organe de l’équilibre, une posture asymétrique est apparue, mais elle n’a pas été corrigée immédiatement lorsque l’autre labyrinthe a été retiré ; par la suite, la posture correspondant à la deuxième opération peut toutefois être adoptée. Ces rapports ont ensuite été vérifiés par Thauer et Peters (124), qui ont confirmé ces résultats et les ont encore différenciés. Curieusement, il s’est avéré que le passage de l’asymétrie à la posture symétrique, lorsqu’il se produisait après une longue période, pouvait se produire par à-coups, c’est-à-dire le même phénomène que celui décrit pour la transplantation musculaire. Une expérience correspondant au nystagmus tournant optocinétique a été réalisée par Gertz. Lorsqu’un sujet tourne sur un plateau tournant de manière à conserver strictement sa position dans l’espace et qu’il descend après environ une minute, il dévie dans le sens de son propre sens de rotation en marchant et ressent une sensation de vertige intense (124a). Sur la question de savoir si les mouvements forcés chez les animaux décérébrés d’un côté sont dus à une particularité de la forme du mouvement ou à une modification accidentelle du tonus des pattes ou autre, Baldus (124b) a procédé à l’ablation de la moitié du cerveau de trois insectes terrestres (Locusta, Dixippus, Carabus) et de deux espèces de coléoptères aquatiques (Dytiscus et Hydrous) ; les animaux ont conservé la tendance au mouvement de manège même lorsqu’ils ont été amputés de pattes du côté opéré. La perte de membres était compensée lors de la marche et de la natation de telle sorte que la direction du mouvement circulaire était toujours maintenue, alors que Loeb avait auparavant estimé que celle-ci reposait uniquement sur une activité accrue des membres du côté opéré. Or, il a été expliqué précédemment qu’aucun réflexe n’est directement déclenché par les labyrinthes au niveau des différents organes, mais que le champ de perception enregistre les sensations des différents organes sensoriels et qu’un réflexe apparaît dans les membres, dans la mesure où les mouvements des membres s’orientent sur le champ de perception. La persistance du mouvement se retrouve également dans ce champ de perception, à savoir le nystagmus ; et l’on a pu montrer avec le nystagmus optocinétique qu’il ne dépend pas uniquement des phénomènes d’inertie du liquide céphalorachidien ; car si, lors de la rotation sur la chaise tournante, le nystagmus se produit également suite à l’inversion de l’idée de rotation selon laquelle le sujet est désormais assis et l’environnement bouge, il s’agit alors d’une inertie du champ de perception. Si l’on navigue longtemps sur un bateau qui oscille fortement et que l’on pose ensuite le pied sur la terre ferme, celle-ci semble parfois continuer à osciller pendant un certain temps ; de même, des phénomènes peuvent encore se produire des jours après, selon lesquels la terre semble soudainement osciller, conformément aux mouvements du bateau. Le phénomène de Kohnstamm (125) a une répercussion similaire. Si l’on appuie par exemple fermement sur son bras contre une résistance immuable et qu’on le relâche ensuite soudainement, de sorte que le bras peut se mouvoir librement et sans contrainte, il se déplace alors comme s’il était soulevé par une force dans la direction prévue auparavant. L’impression subjective est comparable à une poussée d’Archimède. Il en va de même lorsque l’on a porté de lourdes charges pendant un certain temps, par exemple sur les épaules, et que l’on s’en débarrasse, on ressent alors un soulagement semblable à une poussée, qui retombe au bout d’un certain temps et se traduit alors plutôt par le contraire, un relâchement ou une lourdeur des membres. Dans le cas pathologique, le principe de performance et la conception peuvent différer. Ce serait le cas si l’organisation des mouvements ne s’inscrivait pas ou pas entièrement dans l’intention de la volonté ; on trouve un tel tableau clinique dans la chorée ; les mouvements permanents et agités rendent extrêmement difficile ce que von Weizsäcker décrit comme prestation. Subjectivement, on a l’impression que quelque chose d’étranger intervient dans les mouvements, qu’il y a un décalage entre l’objectif volontaire et le mouvement réel. La même chose se retrouve dans la pensée, les pensées peuvent certes se dérouler de manière tout à fait logique, mais un cheminement de pensée clair et déterminé est impossible (126). Tant sur le plan physiologique que pathologique, il faut faire la distinction entre la création du mouvement proprement dit et l’intervention dans la musculature au sens du développement de la force. Von Holst a étudié cette question chez les poissons et a mentionné le dualisme entre la fonction motrice et la fonction rythmique automatique dans la moelle épinière (127). D’un point de vue pathologique, cela se produit surtout en cas de raideur musculaire, de rigorisme musculaire, etc. Quel est donc le rapport entre le système nerveux central et les processus de mouvement ? D’un point de vue fonctionnel et histologique, on peut distinguer différents centres et voies. Les différentes fonctions sont-elles en quelque sorte des intégrations de différents points déterminés ? Tout d’abord, on sait que la perte de parties du cerveau et de centres peut être remplacée, dans une certaine mesure, par d’autres parties. Les fonctions de l’hémisphère gauche peuvent être partiellement prises en charge par l’autre hémisphère. Même la perte du cervelet, lorsqu’elle survient chez de jeunes animaux, peut être compensée au fil du temps. Lorsque des parties individuelles sont extirpées, ce ne sont pas des parties individuelles, des fonctions individuelles ou des muscles ou des parties de membres individuels qui disparaissent, mais la capacité de différenciation est d’abord endommagée. Pour l’innervation commune de grands groupes de muscles, une plus petite quantité de substances de cellules pyramidales suffit que pour la fonction différenciée de parties ou de muscles individuels (128). Dans son « Gestaltkreis », Von Weizsäcker établit la règle selon laquelle plus la lésion est complète, plus l’innervation qui est encore possible est collective (129). Cela semble être valable de manière générale, pour la sensibilité comme pour la motilité. Buytendijk (130) a démontré chez le rat que les troubles de l’intelligence après destruction du cerveau dépendaient de la taille du champ et non pas tant de sa localisation. Pour les lignes de la moelle épinière, la difficulté réside dans le fait que pour les quelque quatre millions de points sensoriels de la peau (à l’exclusion de la tête), qui peuvent être différenciés en termes de qualité de sensation et de signes de localisation, on dispose au mieux d’un demi-million de lignes afférentes dans la moelle cervicale (131). Cela pourrait certes être maîtrisé techniquement, mais les essais ultérieurs font apparaître le principe de conduction comme impossible. Rothmann (132) a coupé les voies pyramidales chez des chiens et des singes. Les petites perturbations qui se produisaient étaient compensées au bout de quelques jours, également en coupant le tractus rubrospinal. Des défaillances plus graves n’apparaissaient que lorsque les deux faisceaux étaient coupés, mais des réparations pouvaient également se produire après un certain temps. Les expériences ont été vérifiées par Probst (133), Schäfer (134) et Homes et May (135). Schäfer a sectionné chez le singe les cordons pyramidaux dans la moelle épinière ou les cordons antérieurs et les cordons latéraux voisins. Conformément aux résultats des autres auteurs, la paralysie en cas de lésion pyramidale n’est ni complète ni durable. Or, ces deux systèmes sont les voies motrices les plus importantes. Chez l’homme, on sait que de telles réparations ne sont pas possibles. La section semi-latérale de la moelle épinière ne provoque pratiquement aucune modification chez les vertébrés inférieurs (136). Chez les animaux supérieurs, N. Weiss (137), 164 a constaté que les chiens marchaient à nouveau bien après trois à quatre semaines ; cette constatation a été vérifiée par Schäfer (138) chez le lion, et par Marshall (139) chez le chat. F. Mott a vérifié ces résultats chez le singe (140). Après une section semi-latérale de la moelle épinière, les troubles disparaissent au bout d’un certain temps, et ce plus rapidement pour les fonctions motrices que pour les fonctions sensitives, en particulier pour les fonctions bilatérales symétriques. Osawa a poussé la section encore plus loin (141). Dans un premier temps, une hémisection de la moelle épinière a été réalisée ; peu de temps après, l’animal pouvait à nouveau marcher normalement et se localiser correctement sur le membre postérieur. La section correspondante a ensuite été effectuée un peu plus bas, du côté opposé, avec le même résultat. Une troisième hémisection a ensuite été pratiquée un peu plus bas sur le premier côté, après quoi la marche est redevenue tout à fait normale ; seule la localisation était un peu plus difficile, bien que toujours possible. Une vérification des résultats a été effectuée par R. Danitsch (142). Il ne s’agit donc pas d’un phénomène de coordination de deux unités indépendantes, comme dans le cas de la section totale de la moelle épinière, mais bien d’une poursuite ou d’une déviation multiple. Chez les échinodermes, on ne peut même pas parler d’un système nerveux développé. Uexküll (143) a pu démontrer qu’un petit morceau de coquille ou de réseau nerveux suffit pour un comportement compliqué et coordonné ; peu importe lequel. Plus un animal est jeune et de rang inférieur, plus les fonctions de son système nerveux sont non spécifiques et plus les réparations fonctionnelles après destruction du tissu nerveux sont faciles. Même si l’on peut trouver certaines localisations dans le système nerveux central qui deviennent de plus en plus spécifiques en montant dans le règne animal et surtout chez l’homme, il n’est pas possible d’émettre une hypothèse selon laquelle les fonctions organiques sont contrôlées à partir de la construction du système nerveux central. La fonction se lie certes de plus en plus à la topographie dans la série ascendante du développement, mais elle n’est en principe pas liée à des structures ou organes spécifiques et peut, en cas de défaillance d’un organe, être prise en charge, quoique dans une mesure variable, par d’autres parties de l’organe. La perte de substance cérébrale n’entraîne pas une détérioration de la performance dans le sens d’une défaillance de certains organes ou de certaines fonctions, mais elle affecte la performance de la différenciation et de la spécialisation. La représentation de certains organes ou membres dans le cerveau, telle qu’elle a été constatée par la stimulation, n’est pas une projection structurelle, mais une performance fonctionnelle. La localisation dans l’organisme est également une performance fonctionnelle qui ne peut pas être expliquée par le principe de la représentation des différentes valeurs de lieu par des voies de conduction strictement isolées. Les expériences de recoupement d’Osawa, au cours desquelles chaque voie doit être commutée deux à quatre fois d’un côté à l’autre sans que la localisation ne soit sensiblement perturbée, en sont la preuve absolue. Du point de vue de l’observation de la pensée, il faut dire que le cerveau lui sert certes d’organe, mais ne la produit pas. L’ordre des pensées, dans la mesure où l’homme ne réfléchit pas, mais saisit activement sa pensée, est fondé sur la logique, une loi purement idéelle, et non sur des dépendances métaboliques. Un homme dont les pensées ne répondent pas à la logique et à l’observation des objets, mais dépendent des conditions organiques du cerveau, etc. doit être qualifié de malade. Si les fonctions, bien que représentées de manière localisée dans le système nerveux central, ne sont pas liées aux structures, mais peuvent varier plus ou moins dans les parties des organes en cas de défaillance de la substance, elles doivent représenter quelque chose d’indépendant en soi, qui n’a besoin des organes que pour sa réalisation. La représentativité mutuelle des organes est limitée par la spécialisation croissante. Jusqu’à présent, il a été prouvé que la perception et le mouvement se rencontrent dans l’âme ; cette âme ne peut pas être expliquée hypothétiquement comme étant causée par des processus organiques, elle doit être saisie par des moyens psychologiques. Alors que l’homme peut en principe disposer librement de ses formes de mouvement, celles-ci sont plus ou moins déterminées chez les animaux par les organes existants, c’est-à-dire que ce n’est qu’avec l’apparition de nouvelles situations organiques, par exemple des mutilations, que de nouvelles formes de mouvement apparaissent, qui n’ont jamais été réalisées auparavant. Elles dépendent certes du système nerveux central en ce qui concerne la différenciation et la dédifférenciation et la référence psychologique (performance), mais ne sont pas constituées par ce dernier. La constatation que les différents processus dans le corps sont régulés et que cette régulation est perturbée en cas de coupure des voies nerveuses a conduit au postulat selon lequel les processus organiques sont contrôlés par le système nerveux. Or, le système nerveux est également un tissu qui est contrôlé de la même manière que le reste. Il ne présente pas de phénomènes qui justifieraient sa position d’exception, il fallait donc à nouveau un deuxième principe qui commande à nouveau le système nerveux. Les théories des réseaux nerveux, qui supposent une réaction commune de l’ensemble du système nerveux, peuvent certes expliquer le fait de la réaction commune, mais pas la capacité de la fonction spéciale. Bethe, qui place les processus nerveux uniquement dans le réseau des neurofibrilles — qui, selon des recherches récentes, se révèle de plus en plus être un produit artificiel (144) (il faut être très prudent dans l’interprétation des résultats, comme la membrane criblée dans l’anneau de laçage de Ranvier s’est révélée être un phénomène optique (145)) — s’est attaqué dès 1897 à la théorie du centre (146) et a élaboré le concept de plasticité du système nerveux. Ce faisant, il se trouve toutefois dans une dépendance totale vis-à-vis des conceptions centralistes. Il a d’abord démontré que la cellule ganglionnaire n’avait aucune importance pour le processus nerveux proprement dit. Chez le crabe Carcinus maenas, le ganglion qui alimente la deuxième antenne a été séparé des autres ganglions ; il ne restait qu’une partie du réseau de fibrilles, appelée « neuropilem ». Chez les animaux opérés, l’antenne a conservé sa posture et son excitabilité normales pendant deux à trois jours. « Les cellules ganglionnaires ne sont donc pas nécessaires aux phénomènes essentiels du système nerveux central ; le tonus, la médiation des réflexes et la sommation des stimuli sont également possibles sans cellules ganglionnaires » (147). Cette constatation correspond bien aux autres examens effectués lors de la traversée de nerfs sur des nerfs vasculaires, etc. La cellule ganglionnaire n’a donc qu’une fonction de nutrition pour les fibres nerveuses proprement dites, c’est-à-dire pour la partie du cerveau qui représente l’organe de la pensée non pas en tant que substance grise, mais en tant que substance blanche — un point de vue déjà défendu par Gall il y a plus de 150 ans. Comme il n’est pas possible de prouver l’existence de véritables centres fixes, mais que l’on s’en tient à l’idée que le cerveau régule les autres processus, Bethe défend la théorie selon laquelle le système nerveux central serait constamment modifié dans sa fonction par la périphérie, la régulation supra-nerveuse étant replacée sans plus attendre dans l’ensemble de l’organisme. L’organisme régulé régulerait son régulateur pour maintenir la bonne fonction. Ce milieu plastique doit représenter le réseau de fibrilles. On a utilisé pour cela l’image du couplage glissant, mais il manque totalement celui qui utilise le couplage de manière judicieuse et qui déploie lui-même l’activité. Outre le fait que le psychique ne peut pas être expliqué de manière organique, cette théorie contient la contradiction de se baser sur une hypothèse qu’elle réfute elle-même. Pour pouvoir évaluer correctement la question des réflexes, etc., il faut encore examiner de plus près le rapport entre le mouvement et la perception. Les mouvements ne sont pas seulement des formes de mouvement, ils sont aussi des performances, c’est-à-dire qu’ils se rapportent à des objets. Pour cela, la perception est nécessaire. Pour la performance, il est strictement vrai que le mouvement n’est possible que sur la base de la perception ; il se forme dans le champ de perception. Un chat ébloui ne saute pas de sa chaise. Marcher, courir, sauter, etc. n’est possible que si le terrain est perçu. Seuls les objets qui sont observés d’une manière ou d’une autre peuvent être saisis. Dans les célèbres expériences de Trendelenburg sur les pigeons, que ce dernier a utilisées pour étayer sa théorie de la réflexion en chaîne, la mobilité des ailes est conservée après la section de tous les nerfs sensibles des ailes, mais les animaux ne peuvent plus voler, bien qu’un battement d’ailes ait lieu de temps en temps. Le vol des oiseaux dépend justement du fait que l’air soit perçu comme un milieu dans lequel le mouvement de vol peut être modelé (148). Les pertes de sensibilité sont gênantes lorsque les objets auxquels se rapportent les prestations ne peuvent plus être perçus. Les perceptions ont cependant un caractère d’orientation. Cela a déjà été expliqué pour l’équilibre ; chez les animaux également, le maintien de l’équilibre n’est pas un processus réflexe, mais une orientation. Les poissons, auxquels on a retiré les arcades, se tiennent à l’incidence de la lumière. Chez les insectes, ce phénomène est bien connu sous le nom de réflexe du dos lumineux (149). L’absence de sensibilité de membres entiers peut également être bien compensée. Plus un ataxique intègre ses mouvements dans son champ visuel, plus ses mouvements deviennent sûrs (150). Les singes et les chats peuvent courir et sauter librement après une séparation de la racine postérieure, qui équivaut pratiquement à une désensibilisation (151). Bickel décrit sur ses chiens, sur lesquels il a également pratiqué une section de la racine postérieure et vérifié l’insensibilité, qu’ils pouvaient bientôt courir et sauter de nouveau de manière coordonnée, mais que la coordination s’effondrait complètement lorsque les labyrinthes auriculaires étaient extirpés (152). Comme nous l’avons vu précédemment, cette opération peut sinon être bien compensée en tant que telle. Les troubles moteurs en cas de troubles de la sensibilité ne résultent pas tant d’une absence de la sensibilité que de son changement de fonction, c’est-à-dire qu’elle ne fournit plus de perceptions exactes. Von Weizsäcker indique expressément que l’ataxie chez Tabes n’a rien à voir avec l’absence de réflexes propres dans certaines circonstances, ceux-ci peuvent également manquer sans ataxie et inversement. On sait également, à partir des troubles du champ visuel, que les déficits du champ visuel sont relativement bien tolérés, mais que de petits restes, surtout s’ils ne fournissent pas des sensations exactes, peuvent empêcher une compensation. La dépendance de la motricité à la sensibilité n’existe cependant pas seulement pour la performance — au sens de von Weizsäcker —, car celle-ci se rapporte à des contenus sensoriels extérieurs à l’homme. Même si un contenu idéal ou psychique doit être exprimé, il est nécessaire qu’il puisse être objectivé dans la perception. Lorsque les sourds-muets apprennent à parler et peuvent palper du bout des doigts la parole sur le larynx d’autrui, ils parviennent assez rapidement à former la parole dans la perception des vibrations de leur propre larynx (153) ; Gutzmann décrit cela comme un mouvement de balancier ; les tentatives n’auraient plus de caractère arbitraire. Le mouvement en tant que phénomène psychologique et les dépendances psychologiques évidentes n’ont en fait pas été pris en compte dans la formation de la théorie des réflexes et des centres. Même dans le cas de lésions de parties motrices du cerveau, cet aspect psychologique est beaucoup trop peu pris en compte. Si des ralentissements de la motricité apparaissent en cas de lésions cérébrales, une observation plus précise indique que l’individu doit seulement dépenser beaucoup plus de volonté intérieure pour les mêmes mouvements, et non pas qu’il se passe autant de choses avec le mouvement lui-même. Le système nerveux oppose une plus grande résistance aux intentions de la volonté. 169 Trendelenburg a bien illustré ce phénomène sur des singes (154). Chez le babouin, la région du bras et de la jambe de l’hémisphère cérébral gauche a été coupée à plat. Par la suite, seule la main gauche a été utilisée pour la préhension. Après l’amputation de la main gauche, il était impossible de distinguer la main droite d’une main normale en l’espace de neuf jours. Après une incision profonde de la même zone du cerveau, la nourriture dans la cage était prise uniquement avec la bouche, mais si elle se trouvait en dehors des grilles, la main était à nouveau utilisée. Il s’agit donc clairement d’une difficulté subjective du mouvement. Si l’on examine maintenant les différents termes de réflexe séparément, on constate que des termes psychologiques sont très souvent utilisés pour développer des idées de conduite. Le « réflexe nociceptif » de Sherrington (155) décrit une situation clairement psychologique. Presque toutes les idées réflexes ont une coloration téléologique. Il s’agit là d’une curiosité particulière, car pour qu’une action ait un but, il faut qu’une personne veuille quelque chose et qu’elle ait une vue d’ensemble de la situation ; les différents processus doivent être organisés à partir de sa connaissance précise de manière à ce que le but souhaité soit également atteint. Une machine qui doit remplir sa fonction a besoin d’une personne assez intelligente pour la concevoir. Pour une représentation téléologique de la nature, il faut pour chaque objet fonctionnel une personne dotée de capacités humaines qui l’a construit. Si la finalité était un processus naturel, on n’aurait pas besoin de construire des machines. Si quelqu’un trouve que l’âme est trop mystique en tant que quelque chose d’indépendant en soi et préfère former des concepts téléologiques réflexes sur la base de représentations de conduite matériellement saisissables, il a alors toujours besoin d’un esprit hypothéqué pour que le corps fonctionne à partir de ses données matérielles sans l’aide d’une âme particulière. En omettant de nommer cet esprit, on ne peut pas empêcher qu’il soit contenu dans le concept. Pour un réflexe de protection, il faut quelqu’un qui l’installe. Si, au lieu des interprétations téléologiques, on parle de processus sensés, alors on suppose que le sens est une force de la nature, mais il faudrait d’abord se pencher sur la question du point de vue de science de la nature. Sous la forme connue, ces représentations donnent toutes l’impression d’une métamorphose de l’ancien concept de force vitale, mais on ne nomme pas son nom. Si l’on applique aux réflexes propres, entre autres, le fait constaté précédemment que la forme du mouvement est quelque chose d’indépendant en soi et qu’elle saisit les différents organes pour les mettre en mouvement, il devrait alors s’agir, dans le cas du réflexe propre, du fait qu’un muscle isolé, lorsqu’il risque de tomber hors du mouvement sous l’effet d’une action extérieure, est à nouveau inclus dans la forme du mouvement avec un déploiement de force particulier. Comme nous le démontrerons plus tard, l’explication morphologique du réflexe propre n’est de toute façon pas défendable. Bethe décrit maintenant une observation simple et caractéristique (74). Lorsque l’on descend une montagne et que l’on pose d’abord les talons, de sorte que les muscles extenseurs sont étirés, le réflexe propre se manifeste de manière classique. Si, en marchant, on accroche la pointe du pied à une racine ou autre — les muscles extenseurs se contractent alors à nouveau —, ce n’est pas le réflexe propre qui se produit, mais la réaction inverse, les muscles extenseurs se relâchent encore plus. On peut observer exactement la même chose chez le chien. Les anciennes expériences de Goltz sur le réflexe d’étirement ipsilatéral (156) sont tout à fait analogues. Lorsqu’un chien touche un support avec ses pattes, il étire son extrémité jusqu’à ce qu’il y ait une résistance ferme. Ceci a été considéré comme un réflexe fixe, conditionné par les conditions de conduction. Mais lorsqu’un chien pose une patte sur une trappe et que celle-ci cède soudainement, il tire immédiatement sur sa patte au lieu de l’étirer par réflexe. Alors qu’habituellement, sur un sol mou, il tend la jambe jusqu’à ce qu’il y ait un sol ferme, dans la neige durcie, il retire en quelques secondes un membre qui s’enfonce s’il n’y a pas de sol ferme. Mais le réflexe de Babinsky, évoqué comme un phénomène de conduction, peut également dépendre de situations psychologiques. En effet, chez certaines personnes, il ne se produit que lorsqu’elles sont couchées sur le dos et s’inverse immédiatement en réaction normale lorsqu’elles sont couchées sur le ventre (157). Les phénomènes contradictoires des réflexes deviennent immédiatement compréhensibles si l’on tient compte de leurs relations psychologiques de prestation et d’organisation. Buytendijk (2) avait déjà attiré l’attention sur le fait que même les réflexes inconditionnels classiques dépendent de la « forme du stimulus ». La dépendance de l’effet du stimulus par rapport à la situation initiale a incité Wilder à élaborer ce qu’il appelle la « loi de la valeur initiale » (158). La dépendance des réflexes propres par rapport à l’activité des antagonistes, constatée par Hoffmann (159) dans ses études très précises, est alors compréhensible sans autres hypothèses. Mais il n’y a pas seulement une imbrication avec la sensorialité en ce qui concerne la performance et la conception, le déploiement de la force en tant que tel est également sous une dépendance correspondante. Cette relation a été désignée par Exner sous le nom de sensomobilité (160) et a fait l’objet d’études supplémentaires depuis (161). Au sens classique, ce terme a été utilisé pour désigner la paralysie de la motricité de muscles particulièrement impliqués dans les fonctions de perception, en cas de défaillance de leurs nerfs sensitifs. Au départ, il s’agissait des muscles du larynx et des lèvres. On connaît la célèbre expérience de Ch. Bell qui, en coupant le nerf infraorbitaire de l’âne, a obtenu une immobilisation des lèvres. Magendi a également vu des paralysies lors de la section du cinquième nerf crânien, lorsque seule la sensibilité était détruite. Exner a vu une paralysie du larynx chez le cheval lorsque le nerf laryngé supérieur était sectionné (162). Pineles a sectionné le nerf infraorbitaire chez le cheval et s’est assuré par stimulation électrique que le nerf facial innervait les lèvres ; malgré cela, on a pu observer une paralysie des lèvres lors de l’alimentation (163).L’œsophage du cheval est paralysé lorsque tous les nerfs sensitifs sont sectionnés (164). Polakow a fait des essais sur le chien. En cas de section du nerf infraorbitaire, la lèvre supérieure est également paralysée, en cas d’anesthésie de la langue par sectionnement des nerfs, la léchouille n’est que faiblement affectée et en cas de cocaïne, elle est fortement affectée (165). Une telle défaillance ne démontre pas un phénomène général, mais indique seulement la classification fonctionnelle des muscles concernés. Les phénomènes qui suivent doivent être considérés tout autrement. L’activité sensorielle pure en tant que telle agit de la manière la plus décisive sur l’activité psychique en général, elle a même un caractère suggestif, car on ne peut que difficilement se soustraire aux différentes impressions sensorielles. Si l’être humain n’est pas stimulé par les sens, il a tendance à s’endormir — à moins qu’il ne développe une activité intérieure propre, par exemple une pensée énergique, qui le maintient malgré tout éveillé. Il y a longtemps, on a pu démontrer qu’un cas de lésion cérébrale ne pouvait plus recevoir de stimuli que par un seul organe sensoriel. Si l’on empêchait l’excitation de cet organe, le patient s’endormait. C’est sur ce phénomène que Pavlov en particulier a fondé sa doctrine, et Bickel (96) a développé toute une théorie sur le fait que tout ce qui est psychique n’est que l’apparence de la réactivité. Tout d’abord, l’homme s’éveille et s’active aux stimuli sensoriels. Arpes et Klemm (166) ont constaté des dépendances correspondantes dans l’attention aux stimuli optiques lors de stimuli acoustiques rythmiques. La mobilité des oiseaux est particulièrement dépendante de l’exposition de la rétine. En cas de lumière du jour claire, les pigeons, par exemple, ont une forte propension à se déplacer ; en cas d’éclairage inférieur à 50 lux, ils deviennent remarquablement immobiles, et en cas de manque total de lumière, ils deviennent apathiques et somnolents (167). Chez l’homme, une stimulation rétinienne adéquate a un effet désinhibiteur sur la psychomotricité ; une mesure quantitative a montré que la musculature volontaire était 10 % plus parate en cas d’exposition à la lumière. En cas d’éclairage du champ complet d’un œil, on constate également chez l’homme une plus grande réactivité du côté concerné (168). Les résultats de Patrick vont probablement dans le même sens (169) : des oiseaux ont été coiffés d’un capuchon de velours percé d’un trou pour le bec, ce qui a entraîné un état de demi-sommeil avec une grande insensibilité aux stimuli extérieurs. Cet état était également maintenu lorsque des trous étaient pratiqués pour les oreilles et les yeux. La respiration s’est toujours révélée être un réactif extrêmement fin pour les processus psychiques. Cet état de fait a été largement exploité dans le cadre d’études psychologiques et physiologiques. Or, c’est précisément dans la respiration que se trouve la frontière entre le psychisme individualisé, conscient et arbitraire et le psychisme agissant selon sa propre loi. On peut particulièrement bien observer ici la différence entre les deux processus, à savoir si l’arbitraire viole le psychique agissant involontairement et conduit à des catastrophes ou si les données de l’involontaire sont acceptées comme un fait et que celui-ci devient alors, dans ses limites, entièrement l’expression des intentions enregistrées dans l’arbitraire. C’est ainsi que le langage devient le moyen d’expression le plus fin de l’âme, mais il faut modeler correctement le langage dans les conditions des besoins respiratoires, sinon une dyspnée survient lors d’un discours prolongé. Les stimuli sensoriels entraînent une accélération de la respiration — les très nombreuses études à ce sujet se trouvent chez Wittkower (170). Christiani avait déjà constaté une accélération de la respiration lors d’impressions lumineuses (171). Afin de rendre les résultats aussi indépendants que possible de toutes les composantes psychiques possibles, de telles études ont également été volontiers entreprises sur des nourrissons (172). Si l’on est soudainement exposé à une forte stimulation par le froid, par exemple en étant mouillé avec de l’eau froide, la stimulation respiratoire peut être si forte qu’une apnée inspiratoire se produit. La même chose peut se produire en cas de forte irritation de la zone du trijumeau, particulièrement sensible à cet égard : lorsque l’on sort la tête d’un train en marche, la respiration est facilement « bloquée ». Ce phénomène est particulièrement prononcé chez les nourrissons : il suffit de souffler légèrement sur leur visage pour que l’arrêt respiratoire inspiratoire se produise. Cela clarifie également la question de savoir pourquoi le nourrisson commence à respirer après la naissance ; jusqu’à présent, cela n’était pas clair du tout, car on peut affirmer avec certitude que le taux d’acide carbonique dans le sang n’est pas responsable, car il peut y avoir un taux d’acide carbonique beaucoup plus élevé dans le sang intra-utérin qu’après l’accouchement, sans pour autant entraîner la respiration. Mais les nouvelles stimulations sensorielles à la naissance stimulent le psychisme et la respiration est déclenchée par le psychisme qui s’installe de manière fonctionnelle. Mais l’activité sensorielle en tant que telle n’est pas la seule à agir. Les différentes qualités sensorielles agissent de manière tout à fait différente. La psychologie des couleurs a mis beaucoup de choses en évidence. Nous ne pouvons ici qu’en mentionner quelques-unes. Tout d’abord, il est bien connu que le rouge est particulièrement excitant ; rencontrer un taureau agressif vêtu de rouge comporte un certain risque. Il existe un travail assez fondamental de Zeylmans van Emmichoven (173) sur l’effet psychologique des couleurs en ce qui concerne le tempo psychique, etc. Féré a maintenant étudié la force de flexion des doigts en présence de différentes couleurs (174), Mezger (175) a trouvé une augmentation du tonus des muscles dans le sens de la rigidité en présence d’une lumière bleue et une augmentation de l’activité mobile en présence d’une lumière rouge ; le tonus est devenu fluctuant, cataleptoïde. Bierens de Haan (176) a mesuré chez des canaris et des souris une activité plus importante sous lumière rouge que sous lumière bleue. Goldstein et Rosenthal (177) ont placé des sujets dans une position telle que leurs bras pouvaient pendre librement, ce qui a révélé une polarité entre la lumière rouge et la lumière verte. Le vert agissait dans le sens d’une flexion, et le rouge obtenait une extension. C’est exactement le geste pour les mesures du tonus, le bleu et le vert agissent dans le sens d’un durcissement et d’une contraction, le rouge incite à l’ouverture et à l’activité. Par l’activité sensorielle, l’âme est amenée à s’activer dans le corps ; pour le début de la respiration après la naissance, cela se présente de la manière suivante : les stimuli sensoriels stimulent la respiration et l’aspiration fonctionnelle de l’âme dans l’organisme. Mais il existe encore un autre phénomène fondamental : le développement de la force dans la musculature en général. On distingue en effet physiologiquement le tonus et la contraction. Dans cette dernière, un fait psychologique comme la forme ou la prestation sont déjà impliqués, s’il ne s’agit pas de spasmes ou de contractures, qui représentent déjà un phénomène pathologique. En tant qu’expression de la relation normale de l’âme avec la musculature, il faut d’abord considérer le tonus et l’étudier dans ses dépendances. Le tonus musculaire normal se relâche d’abord immédiatement lors de la section de la racine postérieure, c’est-à-dire lors de la désensibilisation. Ensuite, on trouve tout simplement le tonus en fonction de la sensibilité (178). Lorsqu’un nerf sensitif est coincé à la suite d’un accident ou d’une blessure de guerre, il se produit très souvent une contracture de la musculature correspondante, qui disparaît immédiatement lorsque le nerf est libéré (179). Le phénomène de Brondgeest, l’atonie totale lors de la section des racines postérieures, est utilisé dans l’opération dite de Förster pour lutter contre les spasmes musculaires. Chez l’animal, lorsque le cerveau est sectionné en dessous du nucleus ruber, la rigidité cérébrale dite de Sherrington se produit ; elle est empêchée par la section des racines postérieures, mais peut être provoquée à nouveau par l’irritation des moignons centraux des racines postérieures (180). Les contractures extrapyramidales et hémiplégiques peuvent également être éliminées par l’opération de Förstersehe (181). Même en cas de stimulation artificielle du cortex cérébral, il n’est jamais possible d’atteindre, après désafférentation, des tensions musculaires d’un degré aussi élevé que dans des conditions normales, ce que l’électromyogramme révèle également sans difficulté (182). Même si le développement de la force est en premier lieu lié aux nerfs moteurs, il dépend intimement de la sensorialité, même si, dans un premier temps, ce n’est que de la manière décrite dans l’expérience du chien dont une jambe a été plâtrée, ce qui a perturbé la coordination. Belmonte et Oddi ont découvert, lors d’expériences avec le bain de cocaïne, la dépendance de l’excitabilité des racines antérieures par rapport aux racines postérieures (183). La performance, l’expression, le mouvement et le développement de la force dépendent, comme toute l’activité psychique en général, de l’activité sensorielle et perceptive. Grâce à la perception, l’homme peut vivre directement les processus de l’environnement s’il s’y engage. Tout mouvement doit à son tour intégrer les lois du monde. Il ne doit cependant pas être comparé à une construction, mais à une création artistique. Celui qui veut atteindre un objet avec une pierre doit vivre intensément dans les conditions de la pesanteur et y intégrer la distance de la cible et la forme de la pierre par rapport à la forme du lancer par son propre mouvement corporel. Celui qui voudrait simplement construire ne toucherait jamais rien. Lorsqu’un joueur de tennis renvoie la balle de la manière souhaitée dans n’importe quelle situation, il doit saisir immédiatement la force et la direction de la balle et, à partir de là, créer en un clin d’œil la nouvelle trajectoire de la balle avec son propre mouvement. La théorie des réflexes en chaîne, qui a été réfutée par ailleurs (184), n’est pas possible si l’on examine les performances de plus près. Psychologiquement, la conception du mouvement se distingue toujours du mouvement mécanisé. Les études de Derwort à ce sujet ont déjà été décrites. Mais il existe aussi des études sur la physiologie du travail ; elles montrent que si le rythme du mouvement n’est pas optimal, s’il est imposé par la machine, l’élan uniforme et la structure du mouvement sont fragmentés et que la précision du mouvement diminue en même temps. En même temps, un rejet affectif s’installe comme expression du fait que l’organisation psychique reste insatisfaite et est violée (185). 176 La perception et le mouvement ne peuvent pas être séparés de la manière dont la physiologie l’a théoriquement fait. Si les formes qui y apparaissent sont des constructions, c’est un cas particulier. Chez l’animal en particulier, elles agissent comme une unité parfaite : la perception et le mouvement sont deux fonctions polaires, mais non séparées. Chez l’animal, cette unité d’action psychique est stimulée de l’extérieur par les sens et de l’intérieur par les pulsions. Les deux ne se séparent que chez l’homme. Comme le souligne Buytendijk (2), l’homme et l’animal se distinguent par le fait que, chez l’homme, la perception et l’action doivent être réunies par une activité propre, ce qui est la seule façon de permettre à la liberté de se développer chez l’homme. Les bases psychologiques de la distinction entre les nerfs moteurs et les nerfs sensitifs en tant que deux fonctions séparées ne sont donc pas du tout données. Sur la base des études menées jusqu’à présent, il est tout à fait possible que le nerf moteur transmette une activité sensorielle qui est absolument nécessaire au développement de la force. von Weizsäcker désigne le fait de l’interaction entre la perception et le mouvement par l’expression symbolique « cercle de gestes ». Ses constatations correspondent certes tout à fait aux recherches entreprises ici. Il reste cependant à clarifier la réalité que l’on doit se représenter sous le terme de « Gestaltkreis », à savoir s’il s’agit d’un principe purement psychologique ou d’excitations circulaires entre les nerfs sensitifs et moteurs. Il se voit sans doute amené à cette dernière conception par les résultats des stimulations nerveuses artificielles. D’après les études menées jusqu’à présent, il n’est donc pas possible d’expliquer les phénomènes psychologiques de manière physiologique, c’est-à-dire par des processus purement matériels. Or, la question reste ouverte de savoir où le psychique intervient dans le matériel-physique et comment les deux côtés sont liés ? Le psycho-spirituel de l’homme est-il lié à l’ensemble de l’organisme ou seulement à un organe, par exemple le système nerveux central ? L’idée la plus courante aujourd’hui est que les deux côtés ne se touchent que dans quelques endroits du cerveau et que les effets de l’âme dans le reste de l’organisme doivent être représentés comme des « excitations nerveuses » matériellement saisissables. Cela signifie que le psychique pourrait être pensé comme un « X » physique dans la résultante des effets des forces dans l’organisme. L’ensemble de l’organisme peut alors être saisi selon des lois physico-chimiques, et seul le cerveau est placé dans un « état d’exception » scientifique, parce qu’il s’y produit des « excitations spontanées » qui ne peuvent plus être saisies de manière causale, mais seulement de manière statistique. Le psychique-spirituel est donc « métaphysique » pour l’organisme et « immanent » uniquement pour certaines parties du cerveau. Une telle théorie n’a pas de valeur explicative scientifique, car tous les problèmes ne sont pas résolus, mais seulement déplacés vers le cerveau. Au début, elle ne servait que les besoins subjectifs, car il est plus confortable de continuer à travailler avec les notions acquises sur l’inorganique que d’en acquérir de nouvelles, qui ne peuvent être ni purement physiques ni purement psychologiques, mais doivent englober les deux. Il s’agit maintenant d’examiner si une telle théorie, sous l’angle de laquelle les nerfs moteurs et sensitifs peuvent être considérés comme des voies de conduction de la manière décrite au début et auxquels peuvent être appliqués les résultats de la physiologie des stimuli nerveux, est possible. Une telle théorie peut alors être appelée « théorie des interactions psychophysiques ». Interaction psychophysique Les résultats de la stimulation nerveuse artificielle peuvent-ils être appliqués aux conditions physiologiques ? De nombreuses études, en particulier les études sur les réflexes propres de Hoffmann (159), semblent plaider en ce sens d’un point de vue chronologique. D’autre part, le mouvement musculaire est également possible sans la présence de nerfs.L’utérus peut fonctionner correctement même en cas de dénervation totale au moment de l’accouchement ; la musculature vasculaire du cordon ombilical et du placenta est également dépourvue de nerfs (186). Il est bien connu que le cœur peut également battre sans nerfs. Dans certaines conditions, une stimulation artificielle permet d’obtenir des effets moteurs qui ne sont certainement pas physiologiques. La réaction connue de Vulpian et Heidenhain sur la langue montre qu’après la section du nerf lingual sur la langue 178, des contractions lentes et spiralées de la langue apparaissent après cinq jours lors de la stimulation du nerf hypoglosse (187). Certains auteurs pensent que ce sont les nerfs vasculaires sensibles qui provoquent cet effet ; mais cet effet peut également être provoqué sur d’autres muscles (188). Lors du croisement expérimental de nerfs moteurs et sensitifs, ce phénomène a donné l’illusion d’une union fonctionnelle. En outre, une discussion variée sur ce que l’on appelle « l’innervation tonique » s’en est suivie. Alors que dans l’expérience nerveuse, la vitesse de conduction est indépendante de la taille du stimulus, les mesures du temps de perception pour les stimuli optiques montrent des rapports tout à fait différents (189). Le temps de perception, c’est-à-dire la durée jusqu’à ce qu’un stimulus lumineux arrivant sur la rétine soit ressenti, dépend de l’attention et de l’intensité du stimulus. Il dépend en outre de la cohérence, un fait psychologique découvert par Weizsäkker pour de tout autres études (190). Hoffmann décrit ici qu’un nouveau stimulus lumineux, qui survient par exemple avant une source lumineuse observée, peut apparaître plus tard pour la perception. Dans le cas de deux stimuli objectivement identiques, qui peuvent par exemple être séparés de 1/20 sec, le premier apparaît après le second. L’ensemble du champ visuel en tant que tel peut se contracter et se dilater lors d’un changement soudain de luminosité. Ce mouvement dit gamma (191) contredit la possibilité que la perception dépende de voies de conduction fixes. Ce phénomène est facile à produire : Si l’on cligne rapidement des yeux, on assiste à un changement rythmique rapide correspondant de rétrécissement et d’extension du champ visuel. Les « voies électriques fluides » de Köhler sont une contradiction dans les termes. Il a été démontré précédemment qu’il n’existe pas de spécificité des énergies sensorielles. En ce qui concerne les seuils de stimulation, la puissance sensorielle dépend de l’objet, mais la qualité sensorielle dépend également de l’objet. Il n’existe pas seulement une constance des tailles visuelles, qui vaut d’ailleurs aussi pour l’animal (192), mais aussi l’étrange constance des couleurs visuelles dans certaines limites. Un objet peut conserver sa propre couleur malgré un éclairage différent. Il faut distinguer, du point de vue de la psychologie des sens, entre les objets colorés et la lumière colorée. On n’en a pas tenu compte parce que les stimuli lumineux qui frappent l’œil sont les mêmes ; les phénomènes différents ont été déclarés subjectifs. 179 Les ombres colorées ou ce que l’on appelle le contraste simultané des couleurs n’apparaissent qu’en présence d’une lumière colorée ; les autres phénomènes de contraste, comme le contraste de flore, y correspondent également. Dans le cas de l’ombre colorée, on a une surface blanche éclairée par une source de lumière blanche et une source de lumière colorée, devant laquelle se trouve un bâton ou un objet similaire servant de projecteur d’ombre. L’une des ombres contre la lumière colorée, qui n’est donc plus éclairée que par une lumière incolore, devrait être blanche, mais elle apparaît dans la couleur du contraste. Déclarer cela subjectif n’est pas envisageable, car si l’on a une surface colorée avec une bande blanche, le contraste n’apparaît pas, bien que les deux fois les mêmes ondes lumineuses ou corpuscules de lumière atteignent l’œil. La couleur se comporte aussi différemment selon qu’elle apparaît de manière fonctionnelle dans la surface du fond ou comme objet dans l’espace. C’est ce qu’a particulièrement étudié D. Katz (193). Cela n’est toutefois pas dû à l’organisation particulière de l’homme, mais vaut également pour les animaux (194). Révész l’a étudié pour les poules, V. Bauer, W. Ewald pour les daphnies et A. Kühn pour les abeilles mellifères, tout comme les illusions d’optique sont également valables pour les animaux (195). Cet exemple doit suffire, bien que les phénomènes puissent encore être développés à l’aide de l’optique physiologique et de la psychologie des couleurs. Le fait qu’il ne s’agisse pas simplement d’une transmission de stimuli, mais d’une perception qualitative, est également démontré par le fait qu’une anesthésie de surfaces sensorielles n’est pas simplement vécue comme une absence de stimuli sensoriels, mais comme un engourdissement. Diehl (196) a anesthésié par cataphorèse et par endosmose la zone de sensation en forme de trait apparaissant lors de la stimulation du condensateur du nerf cutané antéropostérieur, puis l’a de nouveau stimulée. L’effet de stimulation n’a cependant pas été localisé dans la même zone, mais dans ses environs. L’irradiation ne permet pas d’expliquer ce phénomène. Il est certes possible de stimuler de manière efférente un nerf sensitif après l’avoir sectionné et de stimuler également les cellules sensorielles intactes dans les zones sensorielles voisines, comme l’a montré Foerster (197), mais l’anesthésie n’a rien changé à ces conditions. Mais comme la localisation est une performance de représentation, cela peut être interprété de telle sorte que de telles sensations de stimulation sont justement localisées dans la région où les sensations correspondantes sont transmises. 180 La condition préalable à tout principe de conduction est une valeur de localisation exacte des différentes voies, c’est-à-dire que les différentes excitations doivent être projetées sur un schéma en raison de leur conducteur déterminé vers le centre ; l’ordre dans le schéma doit cependant être strictement le même que dans l’ensemble de l’organisme, sinon il doit en résulter une confusion inextricable ! Tout d’abord, différents organes terminaux peuvent être attachés à une seule et même fibre. Ohmori a démontré que les corpuscules tactiles de Meissner et les terminaisons nerveuses libres étaient reliés à une fibre (198). Les nerfs sectionnés peuvent être réunis avec une restauration morphologique et fonctionnelle complète. Les nerfs les plus divers des deux moitiés du corps, des régions entières et des nerfs individuels peuvent être croisés entre eux. Bethe (199) a croisé les deux nerfs sciatiques chez des chiens, ce qui a permis de retrouver une pleine capacité et une coordination normale. Crescenzi (200) a sectionné la racine antérieure d’animaux et a inversé le côté et la hauteur. Selon lui, plusieurs zones radiculaires peuvent être alimentées par un seul groupe de cellules radiculaires. En outre, les plexus brachiaux droit et gauche ont pu être échangés (201). Zypkin (202) met l’accent sur le fait que les nerfs qui innervent les muscles striés peuvent être échangés avec ceux qui innervent les muscles lisses (le nerf iléo-inguinal échangé avec le nerf hypogastrique). Kilvington a relié la racine antérieure des segments lombaires, qui innervent la musculature des membres postérieurs, aux moignons périphériques de la racine antérieure, qui conduisent à la vessie et au rectum et en commandent la motricité (203). Les fibres végétatives peuvent être reliées entre elles et aux nerfs spinaux. Ballance (204) a interverti le nerf facial et le nerf glossopharyngien ; le nerf hypoglosse et le nerf facial ont été intervertis plusieurs fois (205), de même que le nerf facial et le nerf vague (206). Erlanger a permuté les nerfs spinaux et le nerf vague (207). Lors de la section du nerf phrénique, l’extrémité centrale a été reliée au nerf récurrent et l’extrémité périphérique à la partie hypoglosse de l’anse hypoglosse (208). Langley et Anderson ont procédé à des études approfondies, ils ont croisé l’extrémité centrale du 5e nerf cervical avec le sympathique (209), le sympathique cervical avec le nerf lingual périphérique (210), la partie centrale du sympathique avec la chorde tympanique (211) (autre union des différentes extrémités nerveuses chez Langley (212)). Mais les nerfs moteurs peuvent aussi être implantés directement dans la musculature paralysée ; Nikolaev a étudié en détail les conditions pour le larynx (213). Ce procédé est également utilisable en clinique, mais un traitement post-exercice intensif est nécessaire pour obtenir un succès complet (214). Pour plus de littérature sur le croisement de petites régions nerveuses, voir (215). Le lien entre les nerfs sensitifs et les nerfs moteurs, autrefois souvent affirmé, n’a pas pu être confirmé d’un point de vue fonctionnel. Bethe a examiné très précisément les tentatives de régénération sur le plan histologique et a dû constater qu’une liaison morphologique est tout à fait possible, mais que les fibres nerveuses motrices qui se développent ne s’incarnent pas dans les organes sensoriels, mais se dirigent vers la musculature où elles forment les plaques terminales. Ce n’est que si l’on ne trouve pas de muscle ou si la musculature rencontrée est déjà innervée que les fibres dégénèrent à nouveau, comme elles le font normalement lors de la régénération. Le rapport entre les organes terminaux est toujours très spécifique, et les organes terminaux surnuméraires dégénèrent à nouveau après la phase de régénération. Les fibres sensitives qui se développent passent à côté du muscle et recherchent la périphérie (216). Ce comportement est conditionné par les propriétés morphogénétiques (mais il n’est pas possible de le développer ici) et ne permet pas encore de tirer des conclusions fonctionnelles. Un phénomène très étrange, sur lequel nous reviendrons plus tard, se produit lors de la connexion entre le nerf facial et le nerf accessoire dans les mouvements conjoints entre les muscles de la mimique et de l’épaule, qui ne peuvent être surmontés que par un exercice prolongé (214, 217). Sinon, de tels mouvements conjoints ne se produisent généralement pas, mais dans ce cas, ils sont toujours tout à fait constants. La régénération de la sensibilité elle-même a été étudiée par Henry Head (218) dans le cadre d’une auto-expérimentation du nerf cutané antébrachial latéral ; dans la première phase de la récupération, l’image d’une dysesthésie s’est clairement manifestée ; excès embarrassant de sensations, localisation imprécise, perte de la différenciation quantitative et qualitative. Ce n’est qu’au bout de plusieurs mois que le patient a retrouvé ses capacités normales. Il supposait donc l’existence d’un deuxième système nerveux qui se régénérait plus lentement, mais cette opinion a été contredite par les examens à nouveau très minutieux et détaillés de Boeke (219). Si seuls les nerfs moteurs étaient échangés, on pourrait encore imaginer que les impulsions seraient régulées de manière centrale par le contrôle permanent du système sensoriel. Pour corriger les voies sensitives mal reliées, il faudrait en plus une capacité de perception. La spécificité de la perception doit donc en tout cas se produire sur le lieu même de l’activité sensorielle ; la seule possibilité d’interpréter mécaniquement la transmission serait que chaque organe sensoriel émette des excitations spécifiques qui seraient déchiffrées par la médiation de voies non spécifiques par un principe de résonance au centre. Cette hypothèse serait contredite par la plastification progressive de la perception dans l’expérience de Head et par les observations de la psychologie des sens. Mais cette question doit être traitée dans le cadre des expériences de Weiss. La question de la correction de la direction des impulsions motrices trouve sa réponse dans les observations de Weiss. P. Weiß a en effet observé, lors de ses transplantations expérimentales de membres sur des anoures, qu’un membre surnuméraire à côté d’un membre normal — lorsqu’il était alimenté nerveusement — suivait les mouvements des membres normaux très précisément et en temps voulu (220). Chaque impulsion touchait donc le muscle correspondant dans les deux membres. Peu importait les nerfs auxquels le greffon était relié, il suffisait d’un petit nerf qui alimentait ensuite l’ensemble du membre par bourgeonnement d’appel. En raison de la prolifération d’une fibre en plusieurs autres, il devait arriver que la même fibre se répartisse sur des muscles très différents. La géométrie de l’alimentation nerveuse n’avait pas d’importance pour le bon fonctionnement dans les limites, si ce n’est que la zone d’alimentation devait se trouver à l’intérieur des segments normaux du membre. Un membre droit greffé sur le côté gauche se déplaçait en miroir, mais de manière synchrone et homologue avec le membre gauche, une jambe se comportait comme un bras dans la région du bras et inversement. La précision du mouvement conjoint était telle que les réflexes propres peuvent être démontrés. Weiß et Verzar ont fait une découverte naturelle : chez une grenouille, il y avait encore deux membres surnuméraires pour un bras normal ; les réflexes propres de chaque membre pouvaient être démontrés sur les deux autres (221). Les résultats de Weiß ont été vérifiés par différents expérimentateurs (222). Hertwig a même constaté ce phénomène de fonction homologue lorsqu’il a transplanté des membres d’espèces différentes, par exemple le triton sur la salamandre. Par des interventions différentes, Weiß a transplanté des muscles individuels d’un membre sur le dos de l’autre côté de crapauds et les a pourvus de nerfs qui ne servaient certainement qu’à d’autres muscles. Après la cicatrisation, les différents muscles étaient stimulés par des stimuli physiologiques, les muscles transplantés ne se contractaient toujours que lors de l’action du muscle identique. Certains d’entre eux réagissaient encore très légèrement à tous les muscles. Des nerfs différents ont été utilisés pour le même muscle et inversement. Seuls les nerfs des extrémités supérieures ne fonctionnaient pas (223). Cela correspondait tout à fait aux résultats de Detwiler, qui confirmait en soi les données de Weiß, à savoir que l’approvisionnement en nerfs est lié au segment de membre. Il suffit parfois d’une très petite connexion nerveuse avec le segment, même si les autres nerfs proviennent d’autres segments (224). Curieusement, les parties les plus immobiles étaient celles proches du tronc, tandis que la main conservait la plus grande mobilité. L’observation selon laquelle certains éléments nerveux s’effilochaient et alimentaient différents organes correspondait bien à celle d’autres auteurs. Kennedy (225) a sectionné chez des chiens tous les nerfs principaux au-dessus du coude (nerfs musculo-cutané, ulnaire, médian, musculo-spiral) et a réuni les quatre moignons périphériques avec le moignon proximal du nerf musculo-spiral (radial). Une fois la régénération terminée, tous les mouvements étaient coordonnés de la même manière. Weiß et Spitzy (226) ont fait de même avec les nerfs tibiaux et péroniers d’un macaque rhésus (Macacus rhesus), ont fibré les extrémités et ont relié les deux extrémités périphériques au moignon proximal du nerf tibial. Langley et Anderson (212) et Langley et Hashimoto (215) ont démontré qu’une fibre nerveuse peut réellement innerver différents muscles en coupant un nerf moteur qui innerve plusieurs muscles, puis en le régénérant. Il a ensuite été sectionné au-dessus de la cicatrice et la branche conduisant à un muscle a été excitée électriquement, ce qui a entraîné la coexcitation d’autres muscles, alors que la coexcitation n’a pas eu lieu lors de la section en dessous de la cicatrice. Bethe (74) a obtenu, après une suture centrale du nerf sciatique et une nouvelle incision haute, une stimulation de la branche conduisant au nerf gastrocnémien, ainsi que de fortes contractions du muscle tibial antérieur, des extenseurs des orteils et du muscle semi-tendineux. Dans le fonctionnement normal, de telles contractions ou troubles de la coordination ne se sont jamais produits. Ces expériences démontrent donc parfaitement que la stimulation artificielle des nerfs n’est pas identique à la fonction physiologique du nerf, ce que Weiß avait déjà signalé. Au début, nous avons attiré l’attention sur le fait que la stimulation nerveuse artificielle ne peut pas être assimilée à l’action de l’âme dans l’organisme, pour des raisons méthodologiques ; le raisonnement méthodologique et l’expérience expérimentale se recoupent donc. Si l’observation de la performance motrice devait justifier une autre conception du réflexe propre, il s’avère ici aussi que les bases morphologiques invoquées sont tout à fait insignifiantes. Le réflexe propre est un phénomène de prestation et non de conduction ! Afin de pouvoir maintenir le principe d’excitation pour la fonction nerveuse, Weiß a postulé le principe de résonance. Chaque stimulus serait donc accordé en soi, passerait par tous les nerfs en même temps, mais n’aurait d’effet que dans l’organe qui possède des résonateurs accordés. Seule la possession des résonateurs déciderait de la fonction de l’organe. En guise d’Experimentum crucis, Weiß proposa de mesurer le courant d’action lors d’un réflexe non seulement sur le nerf correspondant, mais aussi dans d’autres nerfs ; la preuve serait alors apportée, car tous les nerfs donneraient alors simultanément ce signe d’excitation. Wiersma (227) a réalisé cette proposition, mais a constaté que seul le nerf correspondant présentait un courant d’action, les autres nerfs n’en présentant pas. Bethe (74) objecta dans sa discussion que lors d’une permutation ou d’une autre transplantation de muscles, la fonction correcte ne pouvait pas se produire immédiatement ; selon la théorie de Weiß, des troubles du mouvement devaient alors se produire. Alors que le mouvement conjoint s’est avéré non spécifique au côté (la jambe droite fonctionne du côté gauche comme la jambe gauche), aucun trouble du mouvement n’apparaît en cas de croisement des nerfs des deux côtés (193-195), c’est-à-dire que la jambe droite fonctionne toujours comme membre droit malgré le nerf sciatique gauche, ce qui contredit également le principe de résonance. Si des nerfs surnuméraires se sont maintenant accumulés au point que leur co-mouvement est gênant pour le mouvement normal de l’animal, le greffon cesse très rapidement son activité. Par conséquent, si c’était la possession de résonateurs spécifiques 185 et non la fonction qui était déterminante, cela ne pourrait pas être possible. Pour pouvoir maintenir le principe de résonance, il faudrait encore qu’il y ait une faculté particulière qui s’applique aux organes et qui règle les résonateurs de telle sorte que les différents membres soient correctement impliqués dans le mouvement et la prestation, sinon on entrerait en contradiction avec le fait établi que le mouvement n’est pas la résultante mécanique des mouvements individuels de la musculature. Si la théorie de la résonance n’offre donc aucune possibilité d’explication de ces phénomènes et si l’on voulait maintenir le principe d’excitation, il faudrait que chaque impulsion d’excitation soit présente depuis une capacité de perception particulière jusqu’à l’organe final qui, à chaque bifurcation des fibres, dirige l’impulsion sur le bon chemin. Il faut donc dans tous les cas que l’organe soit perçu pour que l’impulsion puisse y être dirigée. Or, il n’y a qu’un seul nerf disponible pour la capacité de perception, et c’est le nerf moteur. Ainsi, il s’avère d’une part que le nerf dit moteur possède une fonction de perception et trouve par ce biais les organes auxquels se rapporte l’impulsion de la volonté. Cette « fonction » a été exprimée pour la première fois par R. Steiner (228). Le processus sensoriel commence donc directement dans l’organe sensoriel lui-même. Les processus physiques, etc. Les processus du monde extérieur peuvent être suivis jusque dans les organes des sens, en particulier dans l’œil, parce que le monde extérieur est perçu dans l’organe des sens. Le processus psychophysique ne se déroule donc pas seulement dans le système nerveux central, mais déjà dans l’organe récepteur lui-même. Il faut encore examiner de plus près un problème de la transplantation, à savoir qu’un bras peut être considéré comme une jambe et inversement. Lorsqu’un bourgeon de membre est transféré à proximité de l’oreille et qu’il se développe, le membre se déplace de manière synchrone avec les mouvements de la mâchoire, et de manière homologue avec les mouvements des branchies lorsqu’il est proche des branchies. Les expériences ont été réalisées par Detwiler (229) et Nicholas (230). Un membre est pris comme mâchoire dans la zone segmentaire correspondante, ce qui signifie qu’il s’agit du même organe, mais sous une forme différente. La jambe, le bras et la mâchoire sont donc des métamorphoses du même organe, car une quelconque ressemblance extérieure entre la jambe et les branchies ou la mâchoire ne peut en réalité pas être affirmée si facilement. Ce fait que les mâchoires sont des membres métamorphosés a également été souligné par R. Steiner (231). C’est ainsi que s’expliquent les étranges mouvements conjoints lors du croisement du facial et de l’accessoire. Ce phénomène a d’ailleurs servi d’argument principal à Versluys (232) dans ses remarques critiques sur la théorie de la résonance de Weiß. Si les mâchoires — y compris la musculature faciale — sont des membres métamorphosés et si le nerf facial périphérique est alimenté par le nerf accessoire, un nerf de la ceinture scapulaire, la musculature faciale est abordée à partir des segments scapulaires comme musculature du bras. Au départ, les deux groupes de muscles fonctionnent comme un seul et même ensemble. Mais comme le succès du mouvement est perçu de l’extérieur, la prestation de différenciation permet de concevoir le mouvement des deux groupes de manière indépendante. Les autres mouvements conjoints qui se produisent parfois lors de croisements de nerfs devraient encore être étudiés plus précisément en ce qui concerne le principe de métamorphose. Mais la question est la suivante : en quoi consiste la réalité de l’organe pour qu’il puisse être perçu en tant que tel ? Qu’est-ce qui est perçu par le nerf moteur ? Ce ne peut pas être la forme extérieure, ce doit être le principe unitaire qui se développe dans les différentes formes, non pas l’état final de l’organe particulier qui s’impose aux sens, mais ce qui est à la base de cet organe. Il y a en général une grande difficulté dans le concept de métamorphose. Tout d’abord, on considère qu’un organe est caractérisé par la somme de ses propriétés. Si l’organe ou l’organisme entier change de propriétés, si le têtard devient grenouille et la chenille papillon, alors on appelle ce processus métamorphose ; c’est un terme historique pour une continuité matérielle ; un objet est devenu un autre. Mais lorsque deux organes ou deux états se trouvent dans un rapport de métamorphose, l’essence de l’organe ne peut pas consister en la somme des propriétés individuelles, si on les aborde comme des organes identiques. Pour les plantes, Goethe a réalisé la métamorphose du principe de formation ; et ce qu’il avait trouvé par voie idéelle, avec une méthode rigoureuse il est vrai, devait être confirmé par la suite par voie expérimentale : à savoir que les différentes parties de la plante sont des métamorphoses du même organe, la feuille. Le principe de formation a été abondamment étudié en zoologie expérimentale ; en ce qui concerne la formation des organes, on a d’abord un champ dans lequel un organe est formé dans son ensemble. Chaque partie de ce champ, si on la retire, donne l’organe entier, la subdivision en champs différenciés n’est qu’un stade ultérieur du développement (233). On ne peut cependant pas attribuer cette capacité à former des organes à la substance en tant que telle, car si l’on retire le tissu à un stade très précoce lors de régénérations et qu’on le greffe dans d’autres champs, ce tissu est alors saisi par la « puissance » de ce champ. Mais les principes de formation peuvent aussi apparaître indépendamment de l’organe et faire valoir leur propre dynamique — cela a été montré pour le champ de perception et en particulier pour le nystagmus optocinétique —, il apparaît alors dans le mouvement comme un principe indépendant par rapport aux organes, mais qui les saisit. Spemann doit également faire remarquer dans sa « Théorie du développement » (233a) que les faits biologiques ont beaucoup plus de ressemblance avec les concepts psychologiques qu’avec les concepts physiques. Quel est donc le rapport entre l’organe et le principe de formation ? Dans la plante, le principe de formation s’exprime entièrement dans la formation des organes, la plante a également une capacité de régénération presque illimitée. Chez les animaux, on peut dire tout d’abord que la capacité de régénération diminue avec la différenciation morphologique, la capacité de création se révèle d’abord dans le mouvement et ensuite dans le niveau de conscience lui-même en tant que représentation. Fortlage (234) a sans doute été le premier à parler d’une polarité de la vitalité et de la conscience. La configuration ne peut pas être désignée comme un résultat des processus matériels, mais le rapport doit être imaginé sur la base des faits observables de telle sorte que le principe configurateur s’exprime dans la matière et représente l’essence même des organes. Selon le degré de développement et de différenciation, le rapport entre les deux parties doit être tout à fait différent, mais il n’est pas possible de s’y attarder ici. Le nerf moteur perçoit donc le principe organisateur de l’organe, dans la mesure où il s’exprime dans le tissu. Comme différents auteurs ont pu le constater, on peut produire chez les anoures un membre surnuméraire en insérant un moignon de nerf central dans une blessure principale (235). Ce qui est produit pour un organe est totalement indépendant du nerf et n’est conditionné que par le territoire. Guyénot divise maintenant le corps en différentes régions (236). La région de la queue, par exemple, s’étend jusqu’à la dernière vertèbre sacrée. Dans toute la région de l’organe, ce principe d’organe est donc actif, sinon on ne pourrait pas produire de nouveaux organes, et un membre transplanté peut être perçu comme tel si la perception de ce membre peut être réunie à la perception de la région, comme organe en quelque sorte aménagé de la puissance d’organe de toute une région. C’est pourquoi, lors de la transplantation, il suffit d’une faible alimentation nerveuse des segments de membres, car c’est alors que la fusion est possible. Le principe de la forme du membre est, de par sa nature propre, quelque chose d’indépendant en soi, qui ne se tourne que vers le matériel. Si l’on retire l’organe lui-même, il réapparaît sous la forme de ce que l’on appelle un « fantôme », dont il a été abondamment question dans la littérature. L’explication populaire par la stimulation nerveuse n’est pas satisfaisante si l’on examine de plus près la constitution de ses manifestations — en particulier le fait qu’il grandisse ou rapetisse occasionnellement et qu’il se rétracte dans le moignon —, même si les phénomènes pathologiques de stimulation jouent un rôle important dans la manière dont le fantôme est ressenti. Si l’on observe la croissance des nerfs, la différence de nature entre les nerfs moteurs et les nerfs sensitifs s’explique. Les moteurs se rapportent au principe de l’organe, les sensitifs aux processus dans la matière, soit aux processus du monde physique extérieur, soit aux processus matériels dans la mesure où ils représentent l’effet des principes de conception. Or, dans la croissance, les nerfs sensitifs sont totalement dépendants de la périphérie et des organes effectivement présents. Un membre transplanté entraîne les nerfs (237) ; si un membre est amputé, il y a moins de nerfs qui se développent et moins de cellules de la corne postérieure se forment (238) ; la peau participe à la perte des cellules ganglionnaires à hauteur de 60% et la musculature à hauteur de 40% ; les cellules de la corne antérieure motrices ne subissent aucune diminution (239). Si l’on étudie la croissance des fibres nerveuses, on constate qu’en cas d’amputation d’un membre, les nerfs sensitifs se développent proportionnellement moins, c’est-à-dire qu’ils dépendent entièrement de la périphérie ou des zones sensorielles existantes. Les nerfs moteurs, en revanche, se développent jusqu’à la cicatrice sans être affectés et ne dégénèrent qu’ensuite, c’est-à-dire qu’ils ne sont pas déterminés par l’organe matériel, mais s’orientent vers le principe de l’organe. Il s’agit donc du même phénomène morphologique et fonctionnel. Dans ses essais, Weiß se demandait lui-même pourquoi un nerf était encore nécessaire, car le principe de résonance permettrait également une transmission des stimuli sans système nerveux, mais il pensait qu’un nerf était nécessaire pour chaque processus nerveux. Mais il ne s’est pas penché sur la question de savoir ce qu’il fallait entendre par processus nerveux. En effet, sa conception de la résonance annulait déjà la notion initiale selon laquelle un changement d’état matériel se propageait à travers des voies de conduction étroitement décrites — à l’instar de la mèche. Si l’ancienne physiologie nerveuse avait imaginé le câble téléphonique, la télégraphie sans fil a pu être développée à partir de ses représentations. C’est cette question de savoir ce qu’est réellement un processus nerveux que nous allons maintenant examiner. Mais tout d’abord, il convient d’inclure les observations faites sur le nerf moteur. Les résultats décrits ci-dessus lors du plâtrage d’un membre chez le chien (74) ont encore d’autres conséquences. Ainsi, si l’on plâtre un membre, le mouvement et la coordination des autres membres sont gravement perturbés. Si ce membre est désensibilisé, le trouble diminue, mais il est toujours présent. Si, au lieu de désensibiliser, on sectionne les racines antérieures, c’est-à-dire les nerfs moteurs, il n’y a aucune gêne, au contraire, on évite même que la jambe qui pend inutilement ne perturbe. Si la perception du mouvement n’était due qu’aux nerfs sensitifs, la coupure des nerfs moteurs ne devrait pas jouer de rôle, mais en réalité, leur intégrité est une condition préalable ; les nerfs sensitifs ne font que renforcer la perception du mouvement des nerfs moteurs. C’est le même phénomène que la dépendance du développement de la force et du tonus musculaire à la sensibilité décrite précédemment : elle renforce, mais ne constitue pas. Lors de l’estimation du poids, les seuils de stimulation s’adaptent strictement à l’objet, comme nous l’avons déjà décrit. Même en cas d’amputation du bras, elle peut se faire de manière très précise avec une prothèse de fracture acide (mouvement des membres provoqué par des tuyaux musculaires dans le moignon) (240). Mais en cas de troubles purement moteurs tels que l’hypertonie, l’hypotonie, l’ataxie cérébelleuse et les parésies dues à la poliomyélite antérieure, dans lesquels la perception du déploiement de la force est donc perturbée, l’estimation du poids se solde régulièrement par une erreur. L’hypertonie entraîne une sous-estimation et l’hypotonie une surestimation du poids. Mais les maladies musculaires pures, comme la myopathie, conduisent au même résultat (241). Von Weizsäcker (5) décrit dans son Gestaltkreis que les mauvaises innervations de la musculature oculaire entraînent un décalage du champ visuel. Ce phénomène est particulièrement évident au niveau de l’œil. Lorsque nous bougeons nos yeux d’avant en arrière, les objets restent immobiles, c’est-à-dire que le champ visuel est déplacé exactement en fonction du mouvement de l’œil, de sorte que les objets apparaissent toujours à l’endroit où ils se trouvent réellement. Si seuls les nerfs sensitifs percevaient le mouvement, il n’y aurait aucune raison de déplacer le champ visuel de manière trompeuse, même en cas de tension musculaire pathologique. E. Th. v. Brücke (242) décrit dans une auto-observation que lors d’une crise de rhumatisme, la natation dans l’eau était ressentie comme difficile. Même sans expérience, le phénomène de Kohnstamm permet de s’en rendre compte à tout moment : Si l’on porte un poids lourd avec un bras et que l’on compare ensuite un poids léger dans les deux mains, celui-ci semble plus léger dans le bras précédemment chargé (243). Altenburger a étudié des patients sur lesquels l’opération de Foerster — section des racines postérieures (sensibles) — avait dû être effectuée. Immédiatement après l’opération, les mouvements étaient fortement ataxiques. Après quelques semaines, la coordination complète était rétablie, les patients pouvaient alors effectuer des mouvements petits et grands, rapides et lents, sous contrôle optique, sans aucun contrôle préalable. Six à huit semaines après l’opération, les patients pouvaient répéter avec précision, les yeux fermés, des mouvements exécutés sous contrôle optique, selon leur ampleur et leur durée, comme des sujets parfaitement sains (244). Von Frey (245) a mené des recherches sur la sensation de mouvement. Il a endormi la sensibilité superficielle et profonde des doigts par anesthésie périneurale, puis il a fait passer un poids sur un rouleau, les yeux cachés, et a modifié le poids. Le mouvement pouvait néanmoins être répété avec précision. Sur la base de réflexions théoriques, il a rendu le périmysium externe et interne, ou les éléments sensoriels qu’il contient, responsables de ce phénomène, sans le vérifier de manière plus approfondie. Au cours de ses recherches sur la proprioceptivité, Fleisch et ses collaborateurs ont découvert un réflexe respiratoire pour lequel seules des voies motrices semblent être nécessaires. Pour la musculature respiratoire du chien, toutes les voies afférentes ont été interrompues, à savoir les nerfs sympathiques et vagaux, les racines postérieures C 3 à C 8 et la moelle épinière elle-même entre C 8 et Th 1. Seul le nerf phrénique était donc resté intact. Lors de l’activation d’une résistance respiratoire, la fréquence respiratoire augmentait rapidement à chaque fois. Mais cela n’était pas dû à la mécanique musculaire, car ce réflexe n’était pas du tout comparable aux effets d’étirement et de charge de la musculature (246). Il est certes connu que les racines antérieures peuvent être traversées par des fibres sensibles individuelles (247), mais celles-ci ne peuvent avoir qu’un volume minimal ; pour des stimuli respiratoires proprioceptifs, il faudrait cependant plus que des stimuli individuels. S’il ne s’agissait que de l’hypertension de quelques fuseaux musculaires, le réflexe respiratoire devrait se produire en cas de stimulation électrique. Conformément aux résultats précédents, le nerf phrénique doit donc percevoir le mouvement respiratoire lui-même. Les nerfs dits efférents ou moteurs perçoivent ce qui vient de l’intérieur de l’organisme, jusque dans sa configuration corporelle. Il ne faut pas oublier que la matière est quelque chose d’étranger à l’âme, en particulier les conceptions modernes de la matière ne laissent pas beaucoup de place à une telle parenté, et ce n’est que dans la mesure où une intériorité s’exprime physiquement et forme un organe que cet organe peut être trouvé par la volonté. Ce processus échappe certes à la conscience normale, mais il a déjà été montré auparavant qu’il faut distinguer différents niveaux de conscience et que différentes perceptions ou prestations sensorielles ne sont pas du tout remarquées en tant que telles, mais qu’elles ne sont remarquées que par leur effet sur d’autres processus, par exemple le stimulus rotatif intempestif. La conscience de veille proprement dite ne se développe qu’à partir des processus sensoriels qui se déroulent dans le monde extérieur. Les fuseaux musculaires, par exemple, perçoivent également ce qui se passe maintenant en termes d’effets de force physiques externes, c’est-à-dire ce qui est devenu le monde extérieur à partir de processus internes. C’est naturellement différent. Les objets des chémorécepteurs ne sont évidemment pas aussi extérieurs que les objets tactiles et visuels. Les prestations sensorielles des organes des sens externes ne sont pas seulement de simples projections des processus naturels ; par les sens, on peut aussi vivre directement l’âme et l’esprit qui s’expriment dans la matière. Ainsi, les processus intérieurs, par exemple les effets de la volonté, peuvent également être vécus à travers leur expression physique dans la conscience de veille. La volonté est vécue dans la conscience éveillée de la même manière que la perception noire : le contenu de la perception proprement dite n’est pas une sensation sensorielle, mais il est saisi conformément à l’objet par les prestations sensorielles environnantes (247). Les processus internes sont perçus par les nerfs moteurs, mais ils apparaissent à la conscience de veille à travers les activités sensorielles externes. On a rendu les fuseaux musculaires responsables de l’expérience de l’activité de la volonté, mais on a fait remarquer que ce n’était pas la volonté elle-même qui était perçue, mais seulement une apparence, à savoir son résultat sous forme de tension musculaire. Or, il s’agit d’une apparence qui reflète un état de fait réel dans l’image extérieure. Si, en raison d’un processus pathologique, la perception intérieure et l’image extérieure diffèrent, il en résulte des sensations désagréables, par exemple que les poids sont surestimés ou sous-estimés. Pour fonctionner correctement, on est certes tributaire des perceptions internes, car sans elles, une activité organique n’est absolument pas possible ; mais on s’oriente tout de même vers l’activité sensorielle externe ; celle-ci a d’abord — comme nous l’avons décrit précédemment — un effet extraordinairement renforçant, presque suggestif. Tout le monde sait combien il est difficile de réprimer des impressions gênantes pendant le travail, par exemple. L’influence « suggestive » peut être telle que l’on s’abandonne intensément au monde extérieur ou que l’on s’en retire — dans tous les cas, le développement de l’initiative personnelle est entravé. W. R. Hess a montré, dans un très bel exposé sur l’immobilisation des parties douloureuses du corps (248), que la douleur entrave de manière décisive le mouvement et l’expression de la volonté, par exemple. Lorsqu’un chien souffrant d’une fracture d’un membre était pris de tremblements dus au froid, la partie douloureuse était exclue de ces tremblements. Avec un bras douloureux, par exemple, on ne peut tout simplement pas déployer la force adéquate, même si l’on prend sur soi la douleur qui survient. Mais l’activité sensorielle peut également être très fortement affectée. Combien de fois nous arrive-t-il, lorsque nous marchons dans la rue en discutant avec quelqu’un, de ne pas remarquer que des choses importantes se passent autour de nous. Mais si l’on veut être particulièrement attentif à certaines perceptions, on évite volontiers d’autres impressions sensorielles, on ferme par exemple les yeux en écoutant. Giessler (249) caractérise la manière dont le champ visuel peut se rétrécir et s’obscurcir soudainement lors d’une violente détonation. Chez l’hystérique, l’âme peut aussi se retirer partiellement, par exemple en cas de paralysie hystérique ou de cécité. Lorsque les impressions sensorielles extérieures disparaissent, il faut se tourner plus intensément vers les perceptions intérieures, ce qui se produit avec un peu d’entraînement. Cette transition est particulièrement bien illustrée dans une expérience de grenouille de Bickel (96). Si l’on sectionne la moelle épinière d’une grenouille, il y a quand même un mouvement actif des extrémités arrière. Ce mouvement disparaît immédiatement si les racines postérieures sont également sectionnées : une paralysie s’installe alors, car il manque une orientation et une stimulation extérieures. Si la section de la racine postérieure n’a lieu que d’un seul côté, ce côté est tout d’abord également paralysé, la jambe sensible bouge conformément au sens ; avec le temps, la jambe paralysée recommence à bouger jusqu’à ce que le mouvement normal soit à nouveau présent. L’absence d’activité sensorielle est relativement facile à compenser, mais en raison de son caractère suggestif, l’activité sensorielle pathologique entraîne des troubles plus graves et moins faciles à corriger. Si les processus pathologiques sont dus à des processus pathologiques dans certains organes nerveux, il peut être tout à fait justifié d’enlever cet organe. Si, sur la base des hypothèses actuelles, les fonctions pathologiques dans lesquelles le système nerveux est impliqué par l’activité de perception sont considérées comme étant causées par celui-ci, cela conduit parfois à des conséquences thérapeutiques assez inquiétantes. Le système nerveux végétatif est au centre de cette problématique. C’est surtout Stöhr et ses élèves (250) qui soutiennent, sur la base d’études histologiques, la relation syncytiale centrale et périphérique de ce système nerveux. Ce système nerveux commanderait tous les processus de manière centrale, c’est pourquoi toutes les maladies fonctionnelles seraient dues à des modifications pathologiques du système nerveux. C’est ce que l’on tente de démontrer par des examens histologiques approfondis. La théorie du réseau remonte à Gerlach, 1871, Golgi, 1886 et Rogiel 1891. Apàthy pensait alors avoir trouvé l’élément conducteur dans les neurofibrilles représentables par coloration (251). Stöhr affirme maintenant qu’il n’existe pas d’éléments nerveux séparés, le plexus mucosus abritant par exemple dans une voie de conduction plasmodiale commune des éléments nerveux afférents, efférents, sécrétoires et vasomoteurs (252). Selon Plaßmann (253), les fibres végétatives passeraient dans les tissus dans le système cellulaire neurohumoral. Le réticulum dit terminal traverserait tous les tissus de manière plus ou moins diffuse. Dans toutes les maladies fonctionnelles possibles, par exemple comme l’asthme, des examens histologiques précis ont permis de trouver une modification des éléments des cellules nerveuses, comme dans les cellules ganglionnaires ou dans le système cellulaire neurohumoral (254). Il faut cependant être conscient des difficultés qui s’opposent à l’interprétation de ces résultats. Si l’on considère tout d’abord que tous les processus organiques sont commandés de manière centrale, le principe de commande devrait être un principe qui fonctionne par lui-même sans commande. Or, le tissu nerveux est tout autant soumis au métabolisme et aux autres conditions tissulaires que les tissus commandés. Si tous les éléments nerveux sans distinction doivent se terminer par un réseau nerveux, comment peut-on se représenter une conduction différenciée ? Une théorie de la conduction suppose en effet des voies de conduction isolées. Ce n’est même pas une théorie logique ! Le fait que le système nerveux soit intégré dans le contexte hormonal par sa production de substances, par exemple, ne dit rien sur la fonction nerveuse. Cette fonction de substance indépendante de la fonction nerveuse se retrouve par exemple dans le tissu chromaffine de la glande surrénale. Le système cellulaire neurohumoral est également une question d’interprétation des résultats histologiques. Déjà lors de la représentation des fibrilles dans le nerf lui-même, les examens optiques de polarisation de Muralts (255) ont montré qu’elles n’existaient pas in vivo sous la forme observée en coloration ; on peut au mieux parler d’une préformation. En ce qui concerne la représentation des fibrilles dans d’autres tissus, par exemple dans ce que l’on appelle le réticulum terminal, il faut tout d’abord se poser la question suivante : à quelle réalité cela correspond-il dans la vie, et ces structures ont-elles encore quelque chose à voir avec la fonction nerveuse ? Comment décider, en présence de telles images histologiques, où se déroule physiologiquement la fonction nerveuse ? En ce qui concerne les résultats des modifications des éléments nerveux, la critique la plus attentive s’impose également. Si l’on constate des modifications dans les cellules ganglionnaires, par exemple en cas de troubles du métabolisme, il faut alors tenir compte du fait que le nerf est également le siège d’un métabolisme destiné à l’entretien de l’organe lui-même. Or, la cellule ganglionnaire semble servir principalement au métabolisme d’entretien, car comme l’a démontré Bethe (144), elle n’est pas nécessaire à la fonction nerveuse. Si le métabolisme se prolonge en quelque sorte dans le système nerveux et que l’on trouve des modifications correspondantes dans la cellule ganglionnaire lors de processus métaboliques pathologiques, cela n’a absolument rien à voir avec un trouble de la fonction nerveuse. Si celle-ci devait être affectée secondairement et que l’on constate cela dans les résultats, l’interprétation correspondant à la théorie respective, selon laquelle la modification du nerf pourrait être liée à un trouble métabolique fonctionnel, peut conduire à des conclusions thérapeutiques tout à fait erronées. Même si l’on constate une augmentation des éléments des cellules nerveuses, il faut tenir compte des expériences. Comme le décrit en particulier Boeke (256) de manière détaillée, les organes terminaux nerveux sont dans un état d’équilibre avec l’organe lui-même ; les éléments qui apparaissent en excès lors de la régénération dégénèrent alors. Les expériences de Detwiler (231-233) ont montré la dépendance des éléments cellulaires centraux vis-à-vis de la périphérie. Des modifications dans les organes eux-mêmes permettent ensuite aux éléments des cellules nerveuses de se placer secondairement dans une nouvelle position d’équilibre (257). Chez la grenouille, l’extirpation précoce d’une phalange d’un membre perturbe le système nerveux et le cerveau, en particulier le mésencéphale. Lors de la classification et surtout de l’interprétation de la fonction des fibres végétatives, il faut également tenir compte du fait qu’un nerf peut percevoir un processus de sécrétion par la perception dirigée vers l’intérieur. Si le nerf est sectionné, l’activité interne ne peut plus trouver l’organe, et en cas de stimulation nerveuse artificielle, ce processus de sécrétion est stimulé. Ce fait, à savoir que la stimulation nerveuse peut imiter le processus qui n’est en fait que perçu, doit être pris en considération. Mais dans le cas de la glande salivaire, qui a fait l’objet d’une étude approfondie, il s’est avéré que la diversité des modifications qualitatives de la salive — telle qu’elle est la règle dans des conditions naturelles — ne peut pas du tout être obtenue par une stimulation nerveuse électrique (258). D’autre part, les influences psychiques, par exemple les affects, ne provoquent pas seulement une augmentation ou une diminution de la sécrétion, mais aussi une modification de la composition des substances, en particulier de la teneur en rhodanium et en azote, de sorte que Wittkower pense que l’affect doit intervenir directement dans la cellule (259). En effet, les nerfs végétatifs peuvent être croisés entre eux — sympathiques ou parasympathiques — ainsi qu’avec les nerfs spinaux, comme nous l’avons montré précédemment. Une difficulté à reconnaître la nature sensible des nerfs moteurs résulte des résultats des courants d’action. Dans les nerfs dits moteurs, des potentiels d’action centrifuges circulent parfaitement et donc certainement certains états d’excitation. Mais pour pouvoir évaluer leur importance pour la fonction nerveuse, il faut d’abord savoir clairement quel est le rapport entre l’activité de perception et les processus matériels. Mais comment les processus nerveux peuvent-ils être stimulés pour l’activité sensorielle ? Pour les nerfs sensitifs, il existe une incitation physique dans les stimulations des organes sensoriels. Mais les nerfs moteurs ne s’adressent pas du tout à ce qui est maintenant le monde extérieur physique, mais ils perçoivent les processus internes dans la mesure où ceux-ci se tournent vers le monde extérieur. Si les organes terminaux moteurs se trouvent dans le bon contexte, ils ne peuvent pas du tout recevoir une stimulation extérieure dans le même sens que les organes terminaux sensibles. Si l’activité de la volonté agissant dans le métabolisme doit donner la stimulation, il n’est pas concevable que cette stimulation puisse se produire à partir de l’organe terminal, car c’est là que l’activité de la volonté est entièrement absorbée dans la perception. Mais l’endroit du nerf qui sert principalement au métabolisme et non à l’activité de perception, où la volonté peut donc s’exercer sans entrave dans le métabolisme, est la cellule ganglionnaire elle-même, de sorte qu’il serait concevable que l’impulsion de la volonté pour un mouvement s’exerce en même temps dans le métabolisme de la cellule ganglionnaire en tant que générateur d’excitation. Mais c’est alors une question d’interprétation des résultats. La présentation la plus détaillée des études sur le courant d’action est donnée par Schäfer (260). Même pour une théorie de l’excitation, les problèmes ne sont pas tous résolus. Il est tout à fait incompréhensible que le courant d’action des organes qui se produit lors d’une activité sensorielle s’intensifie encore une fois avec la cessation de la stimulation sensorielle, comme cela se produit pour certains organes. On a vu une preuve de la relation entre les courants d’action dans les nerfs et les excitations motrices dans les résultats obtenus sur le nerf phrénique, car des salves de courants d’action apparaissent ici dans le rythme respiratoire (261). Mais c’est justement là que l’on doit émettre les plus fortes réserves, car le pouls et le rythme respiratoire se poursuivent dans des régions qui n’ont rien à voir avec le rythme cardiaque et respiratoire. Dans l’hypothalamus (262), on trouve des courants d’action synchrones avec le pouls et la respiration, de même que dans les nerfs cardiaques (263) ; on a même pu déduire des courants synchrones avec la respiration dans le nerf tibial (264). Tout d’abord, on peut seulement dire que le rythme du pouls et de la respiration peut se prolonger dans d’autres processus, sans que ceux-ci soient nécessairement liés à la mécanique circulatoire ou respiratoire. C’est à un tel prolongement fonctionnel du rythme respiratoire dans le domaine de la dynamique circulatoire que nous pensons. Alors que les ondes de pouls ne se transmettent pas ou presque pas à la pression oculaire, les courbes de pression oculaire peuvent être enregistrées de manière synchrone avec l’inspiration et l’expiration. Cela ne peut pas être compris de manière purement mécanique (265). Rijlant souligne qu’il ne s’agit pas de réflexes, mais d’une propagation centrale, lorsque des impulsions motrices vagales et des excitations sympathiques sont trouvées en synchronisation avec la respiration (266). Quelle est donc la relation entre le système nerveux central et le système nerveux périphérique ? Tout d’abord, les sensations individuelles des éléments sensoriels isolés sont réunies. La différenciation proprement dite est une performance des éléments centraux ; 198 comme nous l’avons déjà montré, la perte de substance cérébrale entraîne une dédifférenciation des différentes fonctions. Qu’en est-il des nerfs moteurs ? La différenciation se manifeste de manière très impressionnante, tout d’abord dans la différence entre le système extrapyramidal et les voies pyramidales. Von Weizsäkker, qui attire particulièrement l’attention sur cette différence, la décrit textuellement ainsi : « Il est important de noter que la région cortico-pyramidale, en tant que condition de la spécification, et la région extrapyramidale-cérébrale, en tant que condition du formalisme du mouvement, signifient certes aussi une localisation, mais en fait non pas celle de différentes performances ou fonctions concrètes, mais celle de différents degrés de réalisation d’une idée dans une substance matérielle. Car la spécification, qui est limitée par des pertes de substance de la région cortico-pyramidale, ne se rapporte pas aux parties ou aux éléments du mouvement, mais aux types d’actions, comme se tenir debout, marcher, sauter, saisir, travailler à la main, écrire, etc. De même que le formalisme matériel ne se réfère pas aux espèces, mais aux éléments spatiaux, temporels et intensifs de chaque type de mouvement, de même le tempo, le trajet, l’ampleur des mouvements nécessaires à ces actions » (267). Il faut donc distinguer le fait que chaque mouvement est une configuration et qu’en tant que tel, il saisit les organes, tandis que l’élément proprement volontaire, l’intentionnel en l’homme, modèle les configurations de telle sorte qu’elles soient incluses dans des actions ou des rapports idéels. Lorsque je saisis un objet, ma volonté établit entre cet objet et les mouvements de mes organes un rapport tel qu’il en résulte l’action. Le mouvement devient l’expression complète de l’intention de la volonté. Ceci a déjà été présenté du point de vue psychologique. La différenciation doit déjà commencer dans la moelle épinière, car les troubles caractéristiques apparaissent déjà en cas de lésions des voies de la moelle épinière. Soit l’intervention correcte des intentions dans les mouvements peut être perturbée, ce qui a déjà été décrit pour la chorée. Ce trouble semble être particulièrement présent dans les maladies des ganglions du tronc, et concerne aussi bien les représentations que les mouvements ; la théorie dite de Berge-Reichard (268) a été développée à ce sujet. Mais il se peut aussi que certaines fonctions ne peuvent pas intervenir dans des mouvements d’organes, bien que le mouvement lui-même soit correctement classé dans les prestations. Un exemple très frappant est celui des paralysies du voile du palais. Pour des raisons purement psychiques, par exemple une névrose d’explosion, la musculature du voile du palais peut être totalement défaillante pour la parole, alors qu’elle remplit correctement son rôle dans la déglutition (269). Le même phénomène peut se produire dans les tumeurs de l’angle pontocérébelleux (270). Il n’est pas possible d’aborder ici d’autres aspects de la localisation du système nerveux central, mais il convient de s’attarder sur un aspect fondamental, car il a fortement étayé les théories élaborées jusqu’à présent. La forme et la configuration apparaissent certes comme des phénomènes psychologiques, mais il faut bien comprendre qu’elles peuvent être un contenu psychique, mais qu’elles ne sont elles-mêmes rien de psychique. On ne peut voir de l’âme que dans ce que von Weizsäcker appelle la prestation, c’est-à-dire l’activité intérieure, l’intention, etc. Si l’on considère les fonctions rendues possibles par la moelle épinière, il s’agit de mouvements et de formes de mouvements, l’âme s’oriente entièrement vers la réalisation de la forme de mouvement elle-même ; en particulier dans ce que l’on appelle le réflexe propre, l’âme n’est active que dans la mesure où elle exprime le mouvement ou la posture par le déploiement de force et où elle intègre ou fait entrer les différents membres. L’âme ne s’exprime pas du tout elle-même, elle sert entièrement la forme. C’est pourquoi les animaux dont le système nerveux central n’a plus qu’une moelle épinière intacte donnent l’impression d’être des automates dans leurs réactions, car rien de typiquement psychique n’apparaît. En outre, il a déjà été signalé à plusieurs reprises qu’il existe différents niveaux de conscience qui peuvent se manifester les uns à côté des autres et les uns après les autres, et pour la moelle épinière, c’est en fait la conscience du sommeil qui s’applique tout particulièrement. Les rapports s’inversent dans le cerveau ; ce n’est qu’à travers cet organe que l’âme peut saisir la forme et la façonner, de sorte qu’elle exprime des contenus psychiques ou qu’elle sert des prestations intentionnelles ; c’est là aussi qu’apparaissent la conscience de veille et l’activité de représentation proprement dite. Dans les régions intermédiaires, hypothalamus, ganglions du tronc, etc., semble se trouver le point de basculement ; c’est là qu’apparaît également la vie émotionnelle, dans la mesure où elle concerne la base nerveuse. La vie émotionnelle a également un état de conscience onirique. En ce qui concerne la localisation en général, il faut tenir compte du fait que le psychique est une entité autonome dont l’organe pour l’activité de perception est le système nerveux-sensoriel. Les fonctions de perception ne se différencient que progressivement sur le plan phylogénétique et ontogénétique. De même, l’attribution de plus en plus stricte des parties du cerveau aux fonctions psychologiques ne se forme que progressivement. On peut alors considérer que la fonction se creuse en quelque sorte dans l’organe et que plus une fonction s’est développée dans l’organe, plus elle est liée à cette localisation. Il n’est donc pas possible d’élaborer une théorie unique, mais les résultats individuels, qui doivent être collectés pour chaque cas, permettent de déterminer très concrètement dans quelle mesure la fonction est liée à un organe. Cela est bien sûr très différent pour chaque organisme et chaque organe, car on sait que chaque personne possède des voies d’association très différentes. Mais il faut être très prudent avec les théories sur la formation d’engrammes. Pour la formation de la mémoire, il est admis que les différentes représentations s’impriment dans la substance des cellules ganglionnaires et sont ainsi mémorisées. Mais on peut toujours constater que l’on reconnaît immédiatement un objet que l’on voit pour la première fois dans une partie du champ visuel, lorsqu’on le voit pour la deuxième fois dans une tout autre partie du champ visuel. Il faudrait alors que deux représentations coïncident toujours comme positivement et négativement, ce qui n’est pas le cas dans la pratique. Si l’on considère les faits expérimentaux, on trouve plutôt une relation avec le métabolisme. Grabensberger a mené des recherches sur la mémoire temporelle des insectes en les dressant à des moments précis de la journée et à des intervalles de temps. Après avoir appris le rythme du temps, il a endormi des termites et des fourmis pendant un certain temps avec du chloroforme et de l’éther ; si les animaux se réveillaient à temps, ils arrivaient quand même à l’heure. Mais si l’on donnait de l’iodothyréoglobuline ou si l’on augmentait la température du nid, ce qui augmentait le métabolisme, les animaux arrivaient en avance ; si l’on réduisait l’euchiningaben ou la température du nid, ils étaient en retard (271). Les mêmes résultats ont été obtenus chez les abeilles (272), puis on a étudié l’effet de l’acide salicylique, du phosphore jaune et de l’arsenic blanc sur les fourmis, et on a obtenu des résultats correspondants (273). Gellhorn a récemment constaté que l’administration d’adrénaline, qui agit également sur le métabolisme, fait réapparaître des réflexes conditionnels inhibés (274). Störring rapporte le cas d’un jeune mécanicien qui, à l’âge de 24 ans, avait subi une intoxication au gaz et ne pouvait se souvenir d’aucune impression pendant plus de deux secondes. Mais il arrivait tout seul et ponctuellement aux repas (275). Au moins pour la mémoire temporelle, il faut dire que l’engramme se fait dans l’activité métabolique et est indépendant du cerveau. Celui-ci n’est nécessaire que si la représentation doit avoir lieu ; si la formation de la représentation n’est pas possible, comme chez le patient cité, la mémoire temporelle peut encore agir directement sur la volonté, l’impulsion d’aller manger se réveille au bon moment. Il faut donc que, parallèlement à la représentation, un contenu s’exprime dans l’organisme, de même que le principe du membre s’exprime dans le membre lors de la croissance, que cette empreinte soit à nouveau saisie comme perception interne et puisse à nouveau venir à la représentation lors du souvenir (276). La différence entre l’homme et l’animal est que l’animal est amené à se souvenir par le monde extérieur, alors que l’homme peut disposer librement de sa mémoire par sa propre activité ; sans cela, le développement de la personnalité ne serait pas du tout possible. Les personnes âgées peuvent encore bien se souvenir d’impressions passées, mais très mal de celles qui ont eu lieu à un âge plus avancé. Plus l’organisme vieillit, plus il perd sa plasticité. Dans le cas de troubles psychiques qui étaient auparavant attribués à des modifications du système nerveux central, on sait qu’il s’est avéré dans de nombreux cas qu’il s’agissait d’un trouble du métabolisme, par exemple dans le cas de la schizophrénie, d’une modification du foie (277). Si cette maladie du foie est corrigée, l’état psychique s’améliore également. Pour comprendre la fonction réelle du système nerveux, il convient d’examiner ses propriétés et son implication dans les processus morphologiques. Lorsqu’on ampute un membre d’un triton et sectionne les nerfs qui l’alimentent, la régénération qui se produit normalement n’a pas lieu (278). Chez le triton, ce n’est pas le système nerveux central qui est responsable, mais plutôt les ganglions qui sont nécessaires à la régénération. Schotté fait appel aux ganglions sympathiques. (279) et Piera Locatelli, sur la base d’études de plus de 800 cas, les ganglions spinaux (280). Lorsqu’un membre est en train de se régénérer, qu’il commence déjà à se différencier et que les nerfs sont ensuite sectionnés, les différenciations se reforment et la régénération s’arrondit à nouveau partout. La croissance végétative elle-même n’est pas perturbée par la perte des nerfs, mais plutôt augmentée. Il a déjà été indiqué précédemment (229, 230 ) que l’implantation d’un moignon nerveux central sous la peau stimule le tissu granuleux pour la formation de membres. Si l’on retire le cerveau des grenouilles têtardes avant leur métamorphose, la métamorphose est gravement perturbée, la régression de la queue et des branchies est inhibée, la percée des membres est entravée et ralentie, de même que les transformations du tube digestif (281). Dans ce contexte, le cerveau antérieur ne joue aucun rôle. Lors des expériences de Wolff sur des tritons avec régénération des membres, il s’est avéré que la destruction de la moelle épinière n’avait aucune influence ; seule l’ablation des ganglions spinaux par prélèvement d’un morceau de la colonne vertébrale empêchait la régénération (282). Neumann a suivi cela du point de vue du développement. À l’aide de défauts centraux, il a pu montrer que le premier développement de la musculature et des nerfs moteurs est sous l’influence des centres. Une fois les muscles formés, ils sont indépendants des centres dans leur existence et leur croissance pendant la période embryonnaire et ne redeviennent trophiquement dépendants que plus tard (283). Si une partie quelconque de la chaîne ganglionnaire est détruite chez la chenille de la fausse teigne, la métamorphose est empêchée. Les perturbations varient quelque peu en fonction des ganglions, mais même si un cocon peut être tissé, la métamorphose ne se produit pas (284). Chez les animaux inférieurs, par exemple comme le ver de terre (285) ou les turbellariés (286), on trouve également de telles dépendances du système nerveux lors de la régénération. Korschelt (287) fournit d’autres informations à ce sujet. Le système nerveux conduit donc en quelque sorte la puissance du membre à la réalisation. Il s’est toujours avéré que peu importe quels nerfs sont présents, la formation du membre dépend du champ respectif. Ainsi, ce qui est présent dans un « champ » en termes de principes de formation encore uniformes et indifférenciés est amené par le système nerveux à prendre forme dans la matière. Avec la formation différenciée, on assiste à un durcissement, la formation est liée à une perte de plasticité. Or, c’est dans le système nerveux central lui-même que la différenciation et la formation sont les plus prononcées, à tel point que même la capacité de régénération est perdue. Ce processus, que l’on peut appeler « impulsion de forme », s’exprime le plus fortement dans le système nerveux lui-même et agit à travers lui dans le reste de l’organisme. Pour le têtard grenouille, le cerveau antérieur est déjà sans importance pour l’organisme, pour le triton, seuls les ganglions sont encore actifs de manière correspondante. Mais cette impulsion de forme se tourne dans le système nerveux central dans une autre direction, à savoir vers l’activité de représentation. Psychologiquement, nous avons montré comment la représentation, dans ce que l’on appelle la préforme, est encore totalement active en soi et en mouvement constant ; avec la différenciation et la formation, cette agitation et cette vivacité sont complètement paralysées, de sorte qu’il en résulte, dans le processus final, une image sans vie qui s’objective psychologiquement et qui n’agit plus sur la volonté. Ce n’est qu’à ce moment-là que des éléments individuels apparaissent, et c’est à partir de là que l’activité de l’intellect peut commencer à combiner et à construire ; avant cela, seule une approche artistique et plastique des représentations est possible, ce que l’on appelle historiquement la conscience mythologique. L’homme n’a donc pas dans sa conscience les réalités des représentations, mais les images qu’elles forment. Dans la pensée logique, la progression des représentations n’est pas déterminée par les lois du métabolisme, mais par des lois idéelles, précisément par la logique. Dans sa conscience, l’homme ne vit pas son cerveau, il n’en remarque rien, mais les objets perçus. Si le cerveau ne détermine pas le déroulement des représentations ni n’intervient en tant que représentation elle-même, alors sa fonction ne peut être que celle d’un miroir. Grâce au reflet du système nerveux, l’homme vit le monde dans sa conscience normale. Du fait que les représentations peuvent être complètement paralysées, de sorte qu’elles ne possèdent plus de dynamique propre, l’homme peut s’orienter librement par rapport à elles. Comme la représentation apparaît comme une simple image, qu’elle n’incite pas l’homme à faire quoi que ce soit et qu’il peut librement s’orienter vers elle dans sa volonté, la possibilité du libre arbitre est physiologiquement fondée. Comme les représentations peuvent être utilisées librement — elles peuvent être associées de manière correcte ou incorrecte à des perceptions —, l’homme peut seulement développer une aspiration à la connaissance, sans laquelle la liberté ne serait pas non plus possible. Comme les représentations ont toutes perdu leur caractère vivant, elles ne peuvent plus avoir d’effet plastique ultérieur dans l’organisme. Comme toute image miroir, elles ont une existence fictive. Le processus de formation de l’engramme doit donc être un processus parallèle. La perception de l’engramme peut alors conduire ultérieurement à la représentation. La représentation du caractère de miroir du système nerveux est particulièrement importante pour évaluer les processus d’excitation rencontrés. Pour un processus de réflexion, il n’est évidemment pas important de savoir de quel côté le miroir est entretenu dans sa fonction. Les processus trouvés ne peuvent pas être considérés comme se répercutant sur l’âme. Cela conduit facilement à des erreurs lorsque l’on trouve des images caractéristiques d’électroencéphalogrammes dans les différentes activités psychiques. D’un autre côté, il doit y avoir une sorte de rupture du miroir dans les cellules ganglionnaires. La synapse n’exerce aucune influence sur l’image elle-même et sur l’activité de représentation proprement dite via les cellules ganglionnaires, mais elles jouent un rôle déterminant pour la prestation de différenciation. Une certaine conception de la synapse peut être tout à fait justifiée. La question du lien entre le corps et âme ne peut pas être résolue de manière univoque, et les théories actuelles ne se sont pas révélées efficaces. Le domaine de la capacité de création proprement dite, que l’on peut également qualifier d’organisation de la forme, est quelque chose d’indépendant en soi, aussi bien pour la perception que pour le mouvement. Cette organisation de la forme se modifie de manière caractéristique selon qu’elle se tourne vers l’organisation matérielle ou vers l’âme. La forme du mouvement devient un déploiement de force dans le matériel et une forme de prestation ou d’expression dans le psychique. La forme de perception déploie d’une part la qualité de la stimulation et est modifiée en objet objectif par l’activité de la volonté qui traverse l’organisme. Cette capacité de création est un processus qui est retenu dans le corps par la formation des organes et qui est à la libre disposition de l’âme elle-même. Le lien entre ce qui est d’âme et de corps doit tout d’abord être pensé de manière différenciée. Pour le système nerveux, on ne peut pas parler d’unité corps-âme, contrairement à l’idée habituelle selon laquelle c’est justement en lui que se trouve la transition. D’une manière prudente, la notion de parallélisme psychophysique serait ici appropriée. Avec la vie de la volonté, l’âme intervient directement dans le métabolisme, de sorte que l’on pourrait ici parler le plus légitimement d’une unité. L’âme de l’homme ne coïncide pas non plus avec les limites de l’organisme ; même si l’homme dépend des bases sensorielles, la volonté, dans le processus de perception et de mouvement, dépasse l’organisme de la manière décrite précédemment. En se livrant à la logique dans la pensée, la volonté dépasse fonctionnellement l’organisme par son entrée dans la sphère idéelle. Si on les spécifie, les deux postulats ont aussi une certaine validité : pas de corps sans âme et pas d’âme sans corps. Or, un corps sans âme n’est certainement que le cadavre et, d’une certaine manière, fonctionnellement, l’homme plongé dans un sommeil profond ou totalement anesthésié, qui n’existe que de manière végétative. Mais l’intervention correcte de l’âme fait partie des fonctions physiologiques. Cela a des conséquences méthodologiques, car on ne peut alors pas faire de la physiologie avec des dispositifs expérimentaux qui excluent le psychique. Les théories élaborées sur cette base sont par conséquent fausses. Si l’on nomme âme l’apparence de l’âme en raison de sa relation avec la corporéité, alors la particularité de cette apparence de l’âme est conditionnée par le corps. Si l’on prend au sérieux la proposition « pas d’âme sans corps », on en conclut que les méthodes psychologiques habituelles ne permettent de saisir qu’une certaine forme de l’âme, mais pas l’âme dans sa pleine spécificité. Pour reconnaître celle-ci, il faudrait examiner ce qui est d’âme dans l’état libre du corps. L’animal et l’homme primitif agissent entièrement à partir de l’instinct, la volonté s’active dans le champ de la perception ; les perceptions ne font qu’orienter, elles ne déterminent pas. Ce n’est que chez l’homme libre que le rapport s’inverse : une représentation est acquise par la pensée. Ce n’est que lorsque celle-ci est à nouveau vivifiée par la propre activité de l’homme, c’est-à-dire lorsqu’elle devient un idéal, qu’elle peut impulser la volonté de son propre chef. Alors qu’auparavant, il n’existe qu’une soi-disant liberté de choix, qui n’est qu’une liberté apparente, ici, une liberté créatrice peut naître de la faculté/du patrimoine d’idée. Et c’est en ce point, où l’idée devient comme vivante, « affective », que réside la justification de la théorie de l’interaction : car l’homme est impressionné psychiquement par ce qu’il vit dans son sentiment, et les contenus qui l’émeuvent intérieurement, comme des affects, agissent sur la corporéité de telle sorte qu’ils ne font en quelque sorte que donner une impulsion ou une modification. Pour pouvoir penser l’homme dans sa physiologie, il est nécessaire de concevoir les différents processus comme une interaction de forces physiques-matérielles et de forces non physiques, psychiques-spirituelles, qui se trouvent dans un rapport différent dans chaque cas particulier. Le psycho-spirituel on peut en fait seulement le penser comme un métaphysique pour le système nerveux. Les difficultés méthodologiques ne peuvent pas non plus être contournées par des théories mécanistes. Remarques et références 1 R. Steiner, Von Seelenrätseln, Berlin 1917, S. 230. 2 F. J. J. Buytendijk, Kritik der Reflextheorie, Verhandlungen der deutschen Gesellschaft für innere Medizin 43 (1931). 3 T. Ziehen, Leitfaden der physiologischen Psychologie in 15 Vorlesungen, 5., teilweise umgearbeitete Auflage, Jena 1908. 4 J. von Kries, Allgemeine Sinnesphysiologie, Leipzig 1923. 5 V. von Weizsäcker, Der Gestaltkreis, Theorie der Einheit von Wahrnehmen und Bewegen, Stuttgart 1950. 6 Chr. von Ehrenfels, Über Gestaltqualitäten, Vierteljahrsschrift für Philosophie 14 (1890). Seitdem: V. Benussi, Archiv für die ges. Psychologie 32 (1914). D. Katz, Gestaltpsychologie, Basel 1944. K. Koffka, Principles of Gestalt Psychology, New York 1935. W. Köhler, Gestaltpsychologie, London 1930. Petermann, Die Köhler-Wertheimer-Koffkasche Gestaltspsychologie, Leipzig 1929. 7 R. Steiner, Die Philosophie der Freiheit, Berlin 1894. 8 Rubin, Visuell wahrgenommene Figuren, Kopenhagen 1921, sonst in fast allen gestaltpsychologischen Schriften. K. Koffka, Psychologie der optischen Wahrnehmung, Handbuch der normalen und pathologischen Physiologie, Bd. 12/2, Berlin 1928. 8a A. Zimmer, Zeitschrift für Sinnesphysiologie 47 (1913), S. 106. 9 M. von Senden, Raum und Gestaltauffassung bei operierten Blindgeborenen vor und nach der Operation, Leipzig 1932. 10 E Halpern, Über Raumwahrnehmung mittels des Wärmesinns, Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere (im folgenden: Pflügers Archiv) 202 (1923), S. 274. 11 D. Katz, Der Aufbau der Tastwelt, Zeitschrift für Psychologie und Physiologie der Sinnesorgane, Abtl. 1 Erg. Bd. 11, Leipzig 1925. 12 S. Dykgraaf, Untersuchung über die Funktion der Seitenorgane der Fische, Zeitschrift für vergleichende Physiologie 20 (1934), S. 162. 13 E. Lippert, Über die Unterschiedsempfindlichkeit bei motorischen Gestaltbildungen des Armes, Neue psychologische Studien, 4. Band, Heft 1 (1928). 14 Kamm, Händigkeit und Variationsstatistik, Klinische Wochenschrift 9 (1930), S. 435. 15 A. Bethe, Münchner medizinische Wochenschrift 52 (1905), S. 1228; Archiv für Psychiatrie 76 (1926), S. 81; Skandinavisches Archiv für Physiologie 49 (1926), S. 91; Pflügers Archiv 224 (1930), S. 793. 16 H. Keller, Die Geschichte meines Lebens; W. Stern, Zeitschrift für angewandte Psychologie 3 (1910), S. 327. 17 D. Katz, Der Aufbau der Tastwelt, Zeitschrift für Psychologie und Physiologie der Sinnesorgane, Abtl. 1 Erg. Bd. 11, S. 192, Leipzig 1925. 18 H. Gutzmann, Verhandlungen der deutschen Otologischen Gesellschaft, Wien 1906; Archiv für Laryngologie und Rhinologie 31; Zeitehrift für klinische Medizin 60 (1907). 19 K. Goldstein, Der Aufbau des Organismus, Einführung in die Biologie unter besonderer Berücksichtigung der Erfahrungen am kranken Menschen, Den Haag 1934. 20 V. Benussi, Zeitschrift für Psychologie 29 (1902); Archiv für die ges. Psychologie 24 (1912), S. 31 und 32 (1914), S. 396. F. Mayer-Hillebrand, Zeitschrift für Psychologie 152 (1942), S. 126. 21 O. Abraham, Psychologische Forschung 4 (1923), S. 1. 22 W. Fuchs, Über Farbenänderungen unter dem Einfluß von Gestaltauffassungen, Bericht über den 7. Kongreß für experimentelle Psychologie in Marburg vom 20. - 23. April 1921. 23 K. Fiedler, Das Schwarz-Weiß Problem, Neue psychologische Studien, 2. Band, Heft 3 (1926). 24 Grundlegende Untersuchungen, daß in einer Gestalt das Ganze und die Teile sich gegenseitig beeinflussen, machte schon j. E. Purkinje auf Veranlassung Goethes. Beiträge zur Kenntnis des Sehens in subjektiver Hinsicht, Prag 1815. 25 E. Lenk, Über die optische Auffassung geometrisch-regelmäßiger Gestalten, Neue psychologische Studien, 1. Band, Heft 4 (1926); C. Schneider, Untersuchungen über die Unterschiedsempfindlichkeit verschieden gegliederter optischer Gestalten, Neue psychologische Studien, 4. Band, Heft 1 (1932), S. 89. 26 A. Derwort, Untersuchungen über den Zeitablauf figürlicher Bewegung, Pflügers Archiv 240 (1938), S. 661. 27 E Schumann, Beiträge zur Analyse der Gesichtswahrnehmung, in: Zeitschrift für Psychologie und Physiologie der Sinnesorgane I in 23 (1900) 1; II in 24 (1900) 1; III in 30 (1902) 241 und 321; IV in 36 (1904) 161. 28 F. Sander, Über räumliche Rhythmik, Experimentelle Untersuchungen über rhythmusartige Reihen- und Gruppenbildungen bei simultanen Gesichtseindrücken, Neue psychologische Studien, 1. Band, Heft 2 (1926). 29 E. Wohlfahrt, Der Auffassungsvorgang von kleinen Gestalten, Ein Beispiel zur Psychologie des Vorgestalterlebnisses, Neue psychologische Studien, 4. Band, Heft 3 (1932). 30 F. Sander, Experimentelle Ergebnisse der Gestaltpsychologie, Bericht über den 10. Kongreß für experimentelle Psychologie in Bonn vom 20. —23. April 1927; F. Sander, Gestaltwerden und Gestaltzerfall, 1939; Gemelli, Archiv für Psychologie 65 (1928); U. Undeutsch, Die Aktualgenese, Scientia (Lancaster) 36 (1942). 31 K. Conrad, Über den Begriff der Vorgestalt, Nervenarzt 21 (1950), S. 38; 209 K. Conrad, Über den Begriff der Vorgestalt und seine Bedeutung für die Hirnpathologie, Nervenarzt 18 (1947), S. 289. 32 R. Steiner, Die Rätsel der Philosophie, Berlin 1914; K. Jaspers, Achsenzeit der Weltgeschichte, Vom Ursprung und Ziel der Geschichte, Zürich 1949. 33 A. Heiß, Zum Problem der isolierenden Abstraktion, Genetisch verglei- chende Studien, Neue psychologische Studien, 4. Band, Heft 2 (1930). 34 W. Köhler, Intelligenzprüfungen an Anthropoiden I, Abhandlungen der Königlich Preußischen Akademie der Wissenschaften, Physikalisch-mathematische Klasse 1 (1917). 35 H. Volkelt, Über die Vorstellungen der Tiere, Ein Beitrag zur Entwicklungspsychologie, Leipzig, Phil. Diss. 13.2.1912. 36 K. Conrad, Über die differentiale und integrale Gestaltfunktion und den Begriff der Protopathie, Nervenarzt 19 (1948), S. 315. 37 Daher lehnt Weizsäcker die Gestaltpsychologie für eine Erklärung der Wahrnehmung ab, siehe Der Gestaltkreis, Stuttgart 1950. 38 Die sogenannte Projektion, siehe W. Metzger, Psychologie, Dresden 1941. 39 V. von Weizsäcker, Der Gestaltkreis (s. Anm. 5); Pflügers Archiv 231 (1933), S. 630. 40 P. Christian, Experimenteller Beitrag zur intermodalen vestibulo-optischen Wechselbeziehung der Sinnesorgane, Pflügers Archiv 243 (1940), S. 370. 41 Dittler, Zeitschrift für Sinnesphysiologie 52 (1921), S. 274; Göthlin, Nov. acta roy. Soc. Sci. Uppsala (1927); Könner, Archiv für Augenheilkunde 93 (1923), S. 130. 42 P. Vogel, Über die Bedingungen des optokinetischen Schwindels, Pflügers Archiv 228 (1931), S. 510. 43 M. Hertz, Zur Physiologie des Formen- und Bewegungssehens, Zeitschrift für vergleichende Physiologie 20 (1934), S. 430. 44 Rudolf Steiner beschreibt, daß die Willenstätigkeit den Organismus übergreift (Näheres wird noch später geschildert), Von Seelenrätseln, Berlin 1917. 45 E. Hering, Grundzüge der Lehre vom Lichtsinn, Berlin 1920. 46 A. Rüssel, Über die Helligkeitskonstanz der Sehdinge, Neue psychologische Studien, 6. Band, Heft 3 (1930). 47 E. Gellhorn und K. Weidling, Pflügers Archiv 208 (1925), S. 361. 48 M. von Frey, Sitzungsberichte der Physikalisch-Medizinischen Gesellschaft zu Würzburg, 1913; Zeitschrift für Biologie 63 (1914), S. 129, 203. 49 M. von Frey, Zeitschrift für Biologie 63 (1914), S. 65. 50 W. Fischel, Zeitschrift für Psychologie 98 (1926), S. 342; F. J. J. Buytendijk, Commentationes Pontifica Academicorum Anno IV, Vol. IV, Nr. 13 (1940). 51 Müller und Schumann, Pflügers Archiv 45; Walter, Zeitschrift für Psycho-210 logiet04 (1927), S. 97. 52 A. Derwort, Zeitschrift für Sinnesphysiologie 70 (1943), S. 135. 53 V. von Weizsäcker, Deutsche Zeitschrift für Nervenheilkunde, Verhandlungen der Gesellschaft der Nervenärzte (1926), S. 270. 54 P. Christian und R. Pax, Zeitschrift für Sinnesphysiologie 70 (1913), S. 197. 55 E. Th. von Brücke, Über Sinnestäuschungen auf Grund eines Mißverhältnisses zwischen Kraftempfindungen und wahrgenommener Leistung, Zeitschrift für Sinnesphysiologie 67 (1932), S. 145. 56 W. Biemüller, Wiedergabe der Gliederanzahl und der Gliederungsform optischer Komplexe, Neue psychologische Studien, 4. Band, Heft 2 (1930); W. Hamilton, Lectures on Metaphysics and Logic I, S. 25, Edinburgh & London 1861; Preyer, Biologische Zeitschrift XIV, 4, S. 357; W. Wundt, Grundriß der Psychologie, 8./9. Aufl., Leipzig 1907/09. 57 E Sander, Räumliche Rhythmik, Neue psychologische Studien (s. Anm. 28). 58 K. Koffka, Psychologie der optischen Wahrnehmung, Handbuch der normalen und pathologischen Physiologie 12/2, Berlin 1928. 59 G. Schwarz, Über konzentrische Gesichtsfeldeinengung bei psychisch Normalen, Neue psychologische Studien, 6. Band, Heft 2 (1930), dort weitere Literatur. 60 A. E. Kornmüller, Die Elemente der nervösen Tätigkeit, Stuttgart 1947, dort auch Zusammenfassung; K. Goldstein, Der Aufbau des Organismus, Den Haag 1934; W. Scheidt, Anthropologie, Hamburg 1948. 61 I. P. Pawlow, Die höchste Nerventätigkeit — das Verhalten von Tieren, München 1926. 62 H. Buchholz, Das Problem der Kontinuität, Neue psychologische Studien, 3. Band, Heft 1 (1927). 63 J. von Uexküll und H. Roesen, Der Wirkraum, Pflügers Archiv 217 (1927), S. 72. 64 K. Gerhards, Nichteuklidische Anschauung und optische Täuschung, Naturwissenschaft 24 (1936), S. 437. 65 E. Th. Reye, Die Geometrie der Lage, Vorträge, 2. verm. Auflage, 1. Abtl. Hannover 1877, 2. (Schluß)-Abtl. Hannover 1880; G. C. C. von Staudt, Geometrie der Lage, Nürnberg 1847; J. Steiner, Ges. Werke 1881; J. V. Poncelet, Traité des proprietés des figures, Paris 1822. 66 G. Backmann, Wachstum und organische Zeit, Leipzig 1943. 67 Siehe auch A. Derwort, Zeitschrift für Sinnesphysiologie 70 (1943), S. 135. 68 P. Christian und V. von Weizsäcker, Über das Sehen figurierter Bewegungen von Lichtpunkten, Zeitschrift für Sinnesphysiologie 30 (1943), S. 30. 69 Stein und V. von Weizsäcker, Deutsche Zeitschrift für Nervenheilkunde 99 (1927); V. von Weizsäcker, Zeitschrift für Sinnesphysiologie 64 (1933), S. 79; Nervenarzt 4 (1931). 211 70 Zusammenfassende Darstellung und Literatur bei Bethe, in Studium Generale, März 1930. 71 F. J. J. Buytendijk, Pflügers Archiv 205 (1924). 72 K. S. Lashley, Science 245 (1931). 73 Wird von Weizsäcker im Gestaltkreis eingehend dargestellt (s. Anm. 5). 74 A. Bethe und E. Fischer, Die Anpassungsfähigkeit (Plastizität) des Nervensystems, Handbuch der normalen und pathologischen Physiologie XV, 2, S. 1045; A. Bethe, Plastizität und Zentrenlehre, ebenda, S. 1175. 75 G. E. Coghill, Proc. nat. Acad. Sci. USA 16 (1930), S. 637; Herrik, ibid. S. 643. G. E. Coghill, Anatomy and the problem of Behaviour, Cambridge 1929. 76 P. Weiß, Ergebnisse der Biologie III, S. 134. 77 G. Brown, Journal of Physiology 49 (1915), S. 208. 78 L. Carmichael, Psychological Review (1921), S. 51. 79 B. H. C. Matthews und S. R. Detwiler, Journ. Exper. Zool. 45 (1926), S. 279. 80 Szymanski, Untersuchung über eine natürliche Reaktionsfähigkeit, Psychologische Forschung 2 (1922), S. 298. 81 F. J. J. Buytendijk, Arch. neerl. physiol. 15 (1930), S. 213; Ven, dasselbe 6 (1921). 82 G. Brown, Handbuch der normalen und pathologischen Physiologie X (1927). 83 W. von Buddenbrock, Biologisches Zentralblatt 44 (1921), S. 41. 84 J. von Uexküll, Zeitschrift für Biologie 46 (1905), S. 1. 85 A. Bethe, Pflügers Archiv 224 (1930), S. 793. 86 A. Bickel, Pflügers Archiv 68, S. 110; E. von Holst, Zeitschrift für eine vergleichende Physiologie 20 (1934), S. 582. 87 H. Spemann, Experimentelle Beiträge zu einer Theorie der Entwicklung, Berlin 1936. 88 Thorner, Pflügers Archiv 230 (1932), S. 1. 89 G. Kriszat, Zeitschrift für anatomische Forschung 28 (1932), S. 47. 90 Slotopolsky, siehe W. Goetsch, Handbuch der normalen und pathologischen Physiologie XIII, S. 264. 91 R. Friedländer, Pflügers Archiv 58 (1894), S. 168; J. Loeb, Vergleichende Gehirnphysiologie, Leipzig 1899, S. 57; R. Janzen, Zoologische Jahrbücher, Abteilung für allgemeine Zoologie und Physiologie der Tiere 50, 51, 31. 92 Eichhorst und Naunyn, Archiv für experimentelle Pathologie 2 (1874), S. 225; ebenfalls ließen sich täuschen: Denten, Quelques recherches sur la régénération fonctionelle et anatomique de la moelle epinière, Inaug. Diss. Bern 1873; Brown-Sequard, Archives de physiologie normale et pathologi- 212 que 411892). 93 Schieferdecker, Virchows Archiv 67 (1876), S. 542. 94 I. Steiner, Die Funktionen des Zentralnervensystems und ihre Phylogenese, 4 Bände, 1885 — 1900. 95 ten Cate, Arch. neerl. Physiol. 18 (1933), S. 15, 497. 96 A. Bickel, Pflügers Archiv 65 (1869), S. 231. 97 Hooker, Proceedings of the Society of experimental Biology and Medicine 28 (1930), S. 99. 98 E. Babak, Pflügers Archiv 93, S. 134. 99 Hooker, Journal of experimental Zoology 55 (1930), S. 23. 100 Hooker, Journal of comparative Neurology 38 (1925), S. 315. 101 Hooker und S. Nicholas, Journal of comparative Neurology 50 (1930), S. 413. 102 A. Bickel, Pflügers Archiv 68, S. 110. 103 A. Bickel, Rev. med. de la Suisse rom. 1 (1897), S. 295. 104 Spatz, Deutsche Zeitschrift für Nervenheilkunde 115 (1930), S. 197; Z. Neur. Ref. 19 (1919), S. 320. 105 Spatz, Nissl-Alzheimers Arbeiten, Erg.Bd. (1921), S. 295. 106 F. Goltz und Freusberg, Pflügers Archiv 9, S. 358. 107 Philippson, L'autonomie et la centralisation dans le système nerveux des animaux, Bruxeles 1905. 108 Snyder, Biolbull. Mar. biol. Labor Woods Hole 7 (1904), S. 280. 109 von Baeyer, Pflügers Archiv 227 (1931), S. 171. 110 Perthes, Beiträge zur klinischen Chirurgie 113 (1928), S. 289; Zentralblatt für Chirurgie 46 (1919), S. 471; Rahnke, Archiv für Orthopädie, Mecha- notherapie und Unfallchirurgie 17 (1920), S. 683; Krause, Zentralblatt für" Chirurgie 47 (1920), S. 884. 111 Schulze-Berge, Deutsche medizinische Wochenschrift 43 (1917), S. 433; Zentralblatt für Chirurgie 44 (1917), S. 551; Mau, Verhandlungen der deutschen orthopädischen Gesellschaft, 1929, S. 236; Spitzy, Zeitschrift für orthopädische Chirurgie 46 (1925), S. 111; Katzenstein, Klinische Wochenschrift 2 (1923), S. 2265; Zentralblatt für Chirurgie 50 (1923), S. 1161. 112 Monzardo, Annals of Ophthalmology 9 (1910), S. 605; A. de Kleyn, Duffer de Borenne, XII. internationaler Physiologen-Kongreß Stockholm (1926), S. 88. 113 Marina, Oblath, Danelon, Annali di neurologia 28 (1910), S. 370; Marina, Deutsche Zeitschrift für Nervenheilkunde 44 (1912), S. 138; Neurologisches Zentralblatt 34 (1915), S. 338. 114 E. Jackson, American Journal of Ophthalmology 6 (1923), S. 117. 115 Bei Exartikulation im Ellenbogengelenk, siehe A. Bethe, Plastizität des Nervensystems (s. Anm. 74). 116 A. Derwort, Untersuchungen über den Zeitablauf figurierter Bewegun- gen, Pflügers Archiv 240 (1938), S. 661. 213 117 A. Bethe, Berichte über die gesamte Physiologie und experimentelle Pharmakologie 32 (1925), S. 686; A. Bethe und Woitas, Pflügers Archiv 224 (1930), S. 831; R. Matthaei, Deutsche Zeitschrift für Nervenheilkunde 115 (1930), S. 232. 118 Siehe Anm. 74, A. Bethe, S. 1083. 119 B. Grzimek, in Studium Generale, 3. Jahrgang, Heft 1 (1950). 120 R. Dubois, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées 8, T. II (1885), . S. 642. 121 A. Bethe, Pflügers Archiv 76 (1899), S, 470. 122 A. Bethe, Z. Neur. 148 (1933), S. 783. 123 R. Ewald, Physiologische Untersuchungen über das Endorgan des Nervus octavus, 1892. 124 R. Thauer und G. Peters, Pflügers Archiv 235 (1935), S. 316. 124a H. Gertz, Acta med. scandinav. 57 (1922), S. 41. 124b K. Baldus, Zeitschrift für vergleichende Physiologie 6 (1927), S. 99. 125 0. Kohnstamm, Neurologisches Zentralblatt (1915), S. 290; R. Matthaei, Pflügers Archiv 202 (1923), S. 89; A. Schwartz et P. Meyer, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées 85 (1921), S. 490; Pinkhof, Nederl. tijdschr. v. geneesk. 65 (1921), S. 437; A. Schwartz, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées 91 (1924), S. 1431; A. Samin, Journ. de physiol. et de patte. gen. 23 (1925), S. 790; A. P. C. Forbes, American Journal of Physiology 78 (1926), S. 81; Arch. intern. de Physiol. 27 (1926), S. 353; R. Bellicioni, Arch. di Fisiol. 28 (1930), S. 269. 126 G. Becker, Allgemeine Zeitschrift für Psychiatrie 114; G. Kienle, Die Chorea Huntingtonfälle von 1900 bis Febr. 1947 aus der Universitätsklinik für Nerven- und Gemütskrankheiten der Eberhard-Karl-Universität Tübingen, Tübingen, Med. F., Dissertation vom 26. Mai 1948. 127 E. von Holst, Pflügers Archiv 237 (1936), S. 356. 128 G. Brown, Handbuch der normalen und pathologischen Physiologie X (1927), S. 418. 129 V. von Weizsäcker, Der Gestaltkreis, 3. Auflage, Stuttgart 1947, S. 50. 130 F. J. J. Buytendijk, Ber. Physiol. 61 (1931), S. 345. 131 V. von Weizsäcker, Der Gestaltkreis, S. 55. 132 Rothmann, Berliner klinische Wochenschrift 39 (1902), S. 376, 404; M.schr. Psychiatrie 16 (1904), S. 589; Berliner klinische Wochenschrift 50, Nr. 12, 13 (1913); Archiv für Physiologie (1907), S. 217. 133 Probst, Archiv für Anatomie (1902), Supplement 147. 134 E. A. Schäfer, Quarterly Journal of experimental Physiology 3 (1910), S. 391. 214 135 G. Holmes und M. A. May, Brain 32 (1909), S. 1. 136 Literatur s. bei O. Langendorf, Nagels Handbuch der Physiologie 4 (1909), S. 378. 137 N. Weiss, Sitzungsbericht der Akademie der Wissenschaften, Wien, Math.-nat. Kl. III 80 (1879), S. 340. 138 E. A. Schäfer, Journal of Physiology 24 XXII, 1899. 139 Marshall, Proceedings of the Royal Society of London 57 (1895), S. 475. 140 F. Mott, Journal of Physiology XII (1891). 141 K. Osawa, Untersuchungen über die Leitungsbahnen im Rückenmark des Hundes, Dissertation, Straßburg 1882. 142 R. Danitsch, Zeitschrift für Biologie 81 (1924), S. 241. 143 J. von Uexküll, Zeitschrift für Biologie 37 (1899), S. 334 und 39 (1900), S. 73. 144 A. von Murait, Die Signalübermittlung im Nerven, Basel 1946. Polarisationsoptisch können die Neurofibrillen bestenfalls als vorgebildet betrachtet werden, keineswegs existieren sie in der färberisch sichtbaren Form. 145 H. Lüthy, Experientis, Vol. VI, Tasc. 10, 15, X (1950). 146 A. Bethe, Biologisches Zentralblatt 18 (1898), S. 843. 147 A. Bethe, Archiv für mikr. Anatomie 50 (1897), S. 625; A. Bethe, Allgemeine Anatomie und Physiologie des Nervensystems, Leipzig 1903. 148 W. Trendelenburg, Archiv für Anatomie und Physiologie (1906), 1; Erg. Physiol. 10 (1910), S. 475. 149 F. Margu und E. Bauer, Zeitschrift für Biologie 103 (1950), S. 253; E. von Holst, Publ. Staz. Zool. Napoli 15 (1935), S. 15. W. von Buddenbrock, Grundriß der vergleichenden Physiologie, 1: Physiologie der Sinnesorgane und des Nervensystems, Berlin 1937, 2: Atmung, Blut, Erfolgsorgane, Blutkreislauf, Berlin 1937. 150 O. Foerster, Restitut. der Motilität, Deutsche Zeitschrift für Nervenheilkunde 115 (1930), S. 248. 151 F. Hartmann und W. Trendelenburg, Zeitschrift für die ges. experimentelle Medizin 50 (1926), S. 280. 152 A. Bickel, Pflügers Archiv 67 (1897), S. 299. 153 Gutzmann, Verhandlungen der Deutschen Otologischen Gesellschaft auf der 15. Versammlung in Wien 1906, Hrsg.: A. Denker; Archiv für Laryngologie und Rhinologie 31 (1918); Zeitschrift für Klinische Medizin 60 (1907). 154 W. Trendelenburg, Zeitschrift für Biologie 65 (1915), S. 103. 155 C. S. Sherrington, The integrative action of nervous System, London 1906. 156 F. Goltz, Pflügers Archiv 20 (1879), S. 1. 157 Mankowsky und W. Bader, Deutsche Zeitschrift für Nervenheilkunde 88 (1926), S. 42. 158 Wilder, Klinische Wochenschrift 10, Nr. 14 (1931). 159 Hoffmann, Die Eigenreflexe, 1922; siehe auch Erg. Physiol. 215 160 S. Exner, Entwurf zu einer physiologischen Erklärung der psychischen Erscheinungen I, S. 6, Leipzig und Wien 1894. 161 A. Kreidl, Die Sensomobilität, Handbuch der Physiologie IX, (1930), S. 761; Foerster, Handbuch der Neurologie 5 (1936), S. 304. 162 S. Exner, Zentralblatt für Physiologie 3 (1890), S. 115. 163 E Pineles, Zentralblatt der Physiologie 4 (1891), S. 741. 164 Chaveau, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées (1891), S. 155. 165 Polakow, Archiv der Psychiatr. Neurologie und ger. Psychopathie, Charkow (1894), H. 2 und 3. 166 G. F. Arpes und O. Klemm, Psychologische Studien IX (1909). 167 Raband und Verrier, Comptes rendus hebdomadaires des séances de l'académie des sciences (Paris) 189 (1934), S. 2280. 168 J. van Essen, Zeitschrift für Sinnesphysiologie 66 (1935), S. 46. 169 D. M. Patrick, British Journal of experimental Biology 4 (1927), S. 322. 170 E. Wittkower, Einfluß der Gemütsbewegungen auf den Körper, Wien 1936. 171 Christiani, Archiv für Anatomie und Physiologie (1880 und 1886). 172 Peiper, Jahrbuch für Kinderheilkunde und physische Erziehung 123 (1929), S. 129; Ergebnisse der inneren Medizin und Kinderheilkunde 40 (1931), S. 1; Monatschrift für Kinderheilkunde 48 (1930), S. 445; 81 (1939), S. 321; Deutsche medizinische Wochenschrift 48 (1940), S. 495; Jahrbuch für Kinderheilkunde und physische Erziehung 104 (1923), S. 195; Monatsschrift für Kinderheilkunde 29 (1924), S. 236; Jahrbuch für Kinderheilkunde und physische Erziehung 117 (1927), S. 350; weiteres Monatsschrift für Kinderheilkunde 81 (1939), S. 330; Canestrini, Über das Sinnesleben der Neugeborenen, Berlin 1913. 173 J. E. Zeylmans van Emmichoven, De werking der Kleuren van het gevoel, Dissertation, Utrecht 1923. 174 C. Féré, Sensation et mouvement, Paris 1900. 175 A. von Mezger, Albrecht von Gräfes Archiv für Ophthalmologie 127 (1931). 176 J. A. Bierens de Haan, Archiv für Physiologie 19 (1924), S. 45. 177 K. Goldstein und Rosenthal, Schw. Archiv für Neurologie und Psychologie XXVI (1930). 178 F. Margut und E. Bauer, Zeitschrift für Biologie 103 (1950), S. 253; Schaefer und Göpfert, Naunyn-Schmiedebergs Archiv 197 (1938), S. 93. 179 O. Förster, Deutsche Zeitschrift für Nervenheilkunde 115; Handbuch der ärztlichen Erfahrungen im Weltkriege 1914/18, Bd. 4, S. 300, Hrsg. Karl Bonhoeffer, Leipzig 1922. 180 Marinescu, Jonescu-Sisest, Kreindler, Académie roumaine, Bukarest 1947; Dusser de Barenne, H. G., Skand. Archiv für Physiologie 43 (1923), 216 S. 1074Kobori Bunya, Okayama l'gakkai Zasshi 40 (1928), S. 1767. 181 F. M. R. Walche, Encephale 20 (1925), S. 73. 182 Hyde and Gellhorn, American Journal of Physiology 156 (1949), S. 311. 183 Belmonte und Oddi, Arch. ital. d. biol. XV (1891), S. 817. 184 E. von Holst, Pflügers Archiv 235 (1934). 185 von Riedel, Arbeitsbewegungen, Leipzig 1925, S. 200. 186 L. Seitz, Deutsche medizinische Wochenschrift 75 (1950), S. 605. 187 Frank, Nothmann, Kirsch, Kaufmann, Pflügers Archiv 197 (1922), S. 270 und 188 (1921), S. 321; H. Schäffer und H. Licht, Naunyn-Schmiedebergs Archiv 115 (1926), S. 180; G. van Rijnberg, Arch. neerl. Ser. III B. II Nr. 3 (1915/1926), S. 496; L. Orbeli und A. Tonkirch, Russkzj. fisiologiceskij zumal 10 (1927), S. 49; A. Ginezinski und L. Orbeli, Russkij. fisiologiceskif zumal 10 (1927), S. 55. 188 E Bremer, und P. Rijlant, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées 190 (1924), S. 985; Hinsey und H. S. Gasser, American Journal of Physiology 92 (1930), S. 679. 189 Literatur siehe K. Vogelsang, Erg. der Physiologie 26 (1928), S. 122. 190 P. Hoffmann, Nervenarzt 19 (1948). 191 Kenkel, Zeitschrift für Psychologie 67 (1948); Lindemann, Psychologische Forschung 2 (1922), S. 5; K. Koffka und Harrower, Psychologische Forschung 13 (1929), S. 55. 192 O. Kroh, W. Götz, Zeitschrift für Psychologie 99 (1926), S. 247. 193 D. Katz, Die Erscheinungsweisen der Farben, Zeitschrift für Psychologie 7 (1911). 194 G. Révész, Zeitschrift für Psychologie und Physiologie der Sinne, I Abt. 88 (1921), S. 130. 195 G. Révész, British Journal of Psychology 14 (1924), S. 387; G. Révész, System der optischen Raumtäuschungen, 1934. 196 E Diehl, Pflügers Archiv 224 (1930), S. 678. 197 O. Foerster, Neurol., Neuropath. Psych. Psychiatr. Festschrift f Rossolino, 1925, S. 145. 198 Ohmori, Zeitschrift für Anatomie und Entwicklungsgeschichte 70 (1924), S. 347. 199 A. Bethe, Münchner Medizinische Wochenschrift 52 (1905). S. 1228. 200 G. Crescenzi, Chirurgia degli organi di movimento (1924), S. 28. 201 W. H. Osborne, und B. Kilvington, Brain 33 (1911), S. 260. 202 J. J. Zypkin, Zur Frage über das Zusammenwachsen der peripheren Nerven und der hinteren Rückenmarkswurzeln, Kasan 1910. 203 B. Kilvington, British Medical Journal 1 (1907), S. 988. 204 C. Ballance, British Journal of Surgery 11 (1923), S. 327; British Medical Journal 2 (1924), S. 349. 205 Koerte, Deutsche Medizinische Wochenschrift 29 (1903), S. 293; A. Wert- 217 heim, Deutsche Zeitschrift für Chirurgie 137 (1916), S. 147; H. F. O. Haberlandt, Zentralblatt für Chirurgie 43 (1916), S. 74; Colledge, British Medical Journal 1 (1925), S. 547. 206 Barrago-Ciarelli, Zit. Zentralblatt für Chirurgie 28 (1901), S. 718. 207 J. Erlanger, American Journal of Physiology 13, S. 372. 208 Colledge und C. Ballance, loc. cit. 1 (1927), S. 553, 609. 209 J. N. Langley und H. K. Anderson, Journal of Physiology 30 (1903), S. 439. 210 J. N. Langley und H. K. Anderson, Proceedings of the Royal Society of London 73, S. 99. 211 J. N. Langley und H. K. Anderson, Archivio di fisiologia 31, S. 365. 212 J. N. Langley und H. K. Anderson, Journal of Physiology 31 (1904), S. 365; J. N. Langley, Proceedings of the Royal Society of London 62 (1898), S. 331. 213 Nikolajew, Monatsschrift für Ohrenheilkunde, Laryngologie und Rhinologie 61 (1927), S. 923, 1005. 214 O. Foerster, Restitution der Motilität, Deutsche Zeitschrift für Nervenheilkunde 115 (1930), S. 248. 215 A. S. J. Blalock, Crowe Archiv of Surgery 12 (1926), S. 95; C. Balance, Colledge, Bailay, British Journal of Surgery 13 (1926), S. 533; G. E. Burget, American Journal of Physiology 44 (1917), S. 492. W. B. Cannon, American Journal of Physiology 36 (1915), S. 362. Calugareanu und V. Henri, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées Paris (1900), S. 503; Journal de physiologie et de pathologie générale 2 (1900), S. 709; R. H. Cunnigham, American Journal of Physiology 1 (1898), S. 293; Gunn, Transactions of the American Surgical Association 4 (1886); Hoessly, Beiträge zur klinischen Chirurgie 99 (1916), S. 186; R. Kennedy, Proceedings of the Royal Society of London, Ser. B. 84, S. 75; ibid. 87 (1913), S. 331;. B. Kilvington, British medical Journal 1 (1905), S. 935; ibid. 2 (1905), S. 625; B. Kilvington und W. A. Osborne, Journal of Physiology 34, S. 267; J. N. Langley und M. Hashimoto, Journal of Physiology 51 (1917), S. 318; D. I. M. Marine, Rogogg und C. N. Stewart, American Journal of Physiology 45 (1917), S. 269; Muslawsky, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées Paris (1902), S. 841; Rawa, Zentralblatt für die medizinischen Wissenschaften (1883), S. 609; Archiv für Anatomie und Physiologie (1885), S. 296; Archiv für Anatomie und Physiologie 31 (1904), S. 365; A. Stefani, ibid. (1886), S. 488; J. A. Lewin, Zur Lehre von der kreuzweisen Zusammenst. der Nerven, Kalan 1903. 216 J. Boeke, Pflügers Archiv 151 (1913), S. 57; A. Bethe, Pflügers Archiv 116 (1907). 218 217 Fauré,#Revue de Chirurgie 18 (1898), S. 1098; P. Manasse, Archiv für klinische Chirurgie 62 (1900), S. 804; A. Cushing, Annals of surgery 37 (1903), S. 641; R. Kennedy, Philosophical Transactions of the Royal Society of London B 194 (1901), S. 127; ibid. 202 (1911), S. 93. 218 H. Head, Studies in Neurology, London 1920. 219 J. Boeke, Handbuch der Neurologie Bd. I, Berlin 1935, S. 995. 220 P. Weiß, Archiv für mikr. Anatomie und Entwicklungsmechanik 102 (1924), S. 635; Ergebnisse der Biologie III (1928), S. 1;Joucnal of comparative neurology 40 (1926), S. 241. 221 F. Verzar und P. Weiß, Pflügers Archiv 223 (1929), S. 671. 222 W. Brandt, Roux Archiv 106 (1925), S. 193; Glimtz, Roux Archiv 106, S. 358; L. Gräper, Archiv für mikr. Anatomie und Entwicklungsmechanik 102 (1924), S. 263; G. Hertwig, Sitzungsberichte und Abhandlungen der naturforschenden Gesellschaft zu Rostock III. Folge/1 (1926). 223 P. Weiß, Pflügers Archiv 226 (1930), S. 600. 224 S. R. Detwiler, The Quarterly Review of Biology 1 (1926), S. 61; Journal of Comp. Neurology 38, Nr. 4 (1925), S. 461. 225 R. Kennedy, Proceedings of the Royal Society of London, Ser. B 87 (1913), S. 331. 226 P. Weiß, Ergebnisse der Biologie III, S. 141; Ergänzung zu 220 P. Weiß, Anat. Anz. Erg. H. (1925), S. 206; Klinische Wochenschrift (1929) II, S. 2174. 227 C. A. G. Wiersma, Archives neerlandaises de physiologie de l'homme et des animaux 16 (1931), S. 337. 228 R. Steiner, Vortrag 11. April 1922, den Haag [GA 82]. 229 S. R. Detwiler, Journal of experimental Zoology 55 (1930), S. 319. 230 S. Nicholas, Proceedings of the Society of experimental Biology and Medicine 26 (1929), S. 729; Archiv für Entwicklungsmechanik der Organismen 118 (1929), S. 78. Journal of experimental Zoology 55, 1. 231 R. Steiner, Die Grundimpulse des weltgeschichtlichen Werdens der Menschheit. Acht Vorträge, gehalten zwischen dem 16. September und 1. Oktober 1922 [GA 216]. 232 J. Versluys, Biologia generalis, Archiv für die allgemeinen Fragen der Lebensforschung 3 (1927), S. 385. 233 W. M. Copenhaver, Journal of Experimental Zoology 43, Nr. 3 (1926), S. 321. 233a H. Spemann, Experimentelle Beiträge zu einer Therapie der Entwicklung, (Springer) Berlin 1936, Nachdruck 1968. 234 C. Fortlage, Acht psychologische Vorträge, Jena 1869. 235 E. Guyénot und O. Schotté, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie et de ses filiales et associées 94 (1926), S. 1050. 236 O. Schotté, Comptes rendus hebdomadaires des séances de la Société de physique et d'histoire naturelle de Genève 43 (1926), S. 126. 219 237 S. R. Detwiler, American Journal of Anatomy 33 (1924), S. 407. 238 S. R. Detwiler, Proceedings of the Society of experimental Biology and Medicine 23 (1926), S. 696; S. R. Detwiler, Journal of experimental Zoology 45 (1926), S. 399. 239 S. R. Detwiler, Journal of experimental Zoology 48 (1927), S. 1. 240 J. Borak, Anzeiger der Akademie der Wissenschaften in Wien, Mathematisch-naturwissenschaftliche Klasse (1920), Nr. 13, S. 147; R. Allers und J. Borak, Medizinische Wochenschrift Wien (1920), S. 1165. 241 Panzel, Deutsche Zeitschrift für Nervenheilkunde 87 (1925), S. 161; Lotmar, Zeitschr. Neurot 24 (1921); K. Goldstein, Deutsche Zeitschrift für Nervenheilkunde 88 (1924), S. 68. 242 E. Th. v. Brücke, Zeitschrift für Sinnesphysiologie 67 (1932), S. 145. 243 R. Matthaei, Klinische Wochenschrift 7 (1928), S. 303. 244 H. Altenburger, Zeitschrift für die gesamte Neurologie und Psychiatrie 129 (1930), S. 313. 245 R. Du Mesnil de Rochemont, Zeitschrift für Biologie 84 (1926), S. 522; M. von Frey, Zeitschrift für Biologie 84 (1926), S. 535; Zeitschrift für die gesamte Neurologie und Psychiatrie 104 (1926). 246 A. Fleisch und Tripod, Pflügers Archiv 240 (1938), S. 676. Petitpierre und A. Fleisch, Comptes rendus de la Societé Suisse de Physiologie Jan. (1942), S. 47; Petitpierre, Helv. Physiol. Acta 2 (1944), S. 53 und S. 71; A. Fleisch, Petitpierre, Helv. Physiol. Acta 2 (1944), S. 235; M. Dolivo, Petitpierre, Helv. Physiol. Acta 6 (1948), S. 81. 247 W. Lehmann, Berliner Klinische Wochenschrift 42 (1924), S. 1895; Berliner Klinische Wochenschrift 51 (1920), S. 1218. 248 W. R. Heß, Pflügers Archiv 203, (1924), S. 539. 249 C. M. Gießler, Archiv für d. ges. Psychologie 45 (1923), S. 282. 250 Ph. Stöhr, jun., Ergebnisse der Anatomie und Entwicklungsgeschichte 35 (1941), S. 135; K. A. Reiser, Zeitschrift für die gesamte Neurologie und Psychiatrie 175 (1943), S. 485; Klinisches Monatsblatt der Augenheilkunde 110 (1944), S. 600. 251 A. E. Kornmüller, Die Elemente der nervösen Tätigkeit, Thieme, Stuttgart 1947. 252 Ph. Stöhr, jun., FIAT (Field Information Agency, Technical) Review of German Science 56 (1947), S. 95. 253 Sunder-Plaßmann, Klinische Wochenschrift 21 (1942), S. 469 und 22 (1943), S. 484; Durchblutungsschäden und ihre Behandlung, Stuttgart 1943. 254 Sunder-Plaßmann, Deutsche Zeitschrift für Chirurgie 210; 252 (1939), S. 1; Ph. Stöhr, jun., Ärztliche Wochenschrift 1 (1946). 255 A. von Muralt, Die Signalübermittlung im Nerven, Basel 1946. 256 J. Boeke, Erg. d. Physiol., Biologischen Chemie und experimentellen Pharmakologie 19 (1921), S. 447. 220 257 B. Düyken, Zeitschrift für wissenschaftliche Zoologie 99 (1912), S. 189, Extremitätenexstirpation und Gehirnstörungen. 258 H. Rein., Physiologie des Menschen, 5. und 6. Auflage 1941, S. 195; Babkin, Die äußere Sekretion der Verdauungsdrüsen, 2. Auflage, Berlin 1928; H. Rosemann, Handbuch der norm. und pathologischen Physiologie Bd. III, 1927. 259 E. Wittkower, Einfluß der Gemütsbewegungen auf den Körper, Wien 1936. 260 H. Schaefer, Elektrophysiologie, Bd. I und II, Wien 1940/1942. 261 E. D. Adrian und Bronk, Journal of Physiology 66 (1928), S. 81 und 67 (1929), S. 119; Cardin, Boll. ital. 13 (1938), S. 758; Dittler, Pflügers Archiv 131 (1910), S. 581; 136, S. 533; H. S. Gasser, American Journal of Physiology 85 (1928), S. 372 und 569; W. R. Heß und Wyss, Pflügers Archiv 237 (1936), S. 761, Wyss, Pflügers Archiv 241 (1939), S. 525 und 244 (1941), S. 712; III. Kongr. neurot. internat. (1939), S. 495. 262 R. W. Gerard, W. H. Marshall und Saul, Proceedings of the Society of Experimental Biology and Medicine 109 (1934), S. 38; American Journal of Physiology 30 (1933), S. 1123. 263 E. D. Adrian und Bronk, Journal of Physiology 74 (1932), S. 115; Bronk, Ferguson und Solandt, Proceedings of the Society of experimental Biology and Medicine 31 (1932), S. 579; Govaerts, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 122 (1936), S. 449; P. Rijlant, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 112 (1933), S. 1125. P. Rijlant, Aldayn und Abbelaus, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 122 (1936), S. 791. A. Amann und H. Schaefer, Pflügers Archiv 246 (1943), S. 757. 264 H. Schaefer, Elektrophysiologie, Bd. II, S. 248, Wien 1942. 265 R. Roos, Archiv für Kreislaufforschung 16 (1950), S. 18. 266 P. Rijlant, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 112 (1933), S. 1229. 267 V. von Weizsäcker, Der Gestaltkreis, 3. Auflage, Stuttgart 1947, S. 51. 268 H. Steck, Schweizer Archiv für Neurologie und Psychiatrie 11 (1922), S. 208. 269 0. Muck, Münchner medizinische Wochenschrift 67 Nr. 18 (1920), S. 503. 270 H. W. Stenvers, Nederlandsen tijdschrift voor geneeskunde 65 (1921), S. 637. 271 W. Grabensberger, Zschr. für vergleichende Physiologie 20 (1934), S. 1. 272 W. Grabensberger, Zschr. für vergleichende Physiologie 20 (1934), S. 338.. 273 W. Grabensberger, Zschr. für vergleichende Physiologie 20 (1934), S. 501. 274 E. Gellhorn, Proceedings of the Society of experimental Biology and Medicine 64 (1947), S. 375. 275 G. E. Störring, Über den ersten reinen Fall eines Menschen mit völligem Verlust der Merkfähigkeit, Leipzig 1931. 221 276 H. Poppelbaum, Im Kampf um ein neues Bewußtsein, Dresden 1935, Kap. IV; R. Steiner, Anthroposophie und akademische Wissenschaften 1950, S. 114. 277 R. Fischer, E Georgie, R. Weber und R. M. Piaget, Psychophysische Korrelationen, Schweizer medizinische Wochenschrift 80 (1950), S. 129, dort weitere Literatur. 278 P. Weiß, Archiv für mikroskopische Anatomie und Entwicklungsmechanik 104 (1925), S. 317; Berichte über die gesamte Physiologie und experimentelle Pharmakologie (Hrsg.: Carl Oppenheimer und Peter Rona) 18 (1923), S. 37; 19 (1923), S. 538; 22 (1924), S. 29. 279 O. Schotté, Comptes rendus hebdomadaires des séances de la Société de physique et d'histoire naturelle de Genève; Guyénot und O. Schotté, Comptes rendus hebdomadaires des séances de la Société de physique et d'histoire naturelle de Genève 43 (1926), S. 32; 40 (1923), S. 160; O. Schotte, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 94 (1926), S. 1128; E. Guyénot, ibid. 94 (1926), S. 437. 280 Locatelli, Piera, Archives italiennes de biologie 74 (1924), S. 85; Archivo di science biologiche 7 (1925), S. 301; Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 95 (1926), S. 3. 281 E. Babak, Pflügers Archiv 109 (1905), S. 78. 282 G. Wolff, Archiv für pathologische Anatomie und Physiologie und für klinische Medizin 169 (1902), S. 308. 283 E. Neumann, Archiv für Entwicklungsmechanik der Organismen (1901), S. 448. 284 Metalnikow, Comptes rendus hebdomadaires des séances et mémoires de la Société de Biologie 97 (1927), S. 126. 285 Morgan, Archiv für Entwicklungsmechanik der Organismen 14 (1902). 286 J. M. D. Olmsted, Journal of experimental Zoology 36 (1922). 287 E. Korschelt, Regeneration und Transplantation Bd. I, Berlin 1927, S. 594. Glossaire (brut de traduction automatique) Acide salicylique : acide organique provenant de l'écorce des saules (Salix) Ganglion de l'œsophage : ganglion de cellules nerveuses dans la région de l'œsophage adrénaline : hormone sécrétée par les glandes surrénales affectif : lié à une émotion Anesthésie périneurale : suppression de la sensibilité et de la douleur par l'um-228 Anoures : amphibiens sans queue comme les grenouilles, les crapauds, les crapauds communs Atacticisme : personne dont les mouvements vers un but sont perturbés et désordonnés. Ansa hypoglossi : site du nerf hypoglosse (voir là-bas), caractérisé par l'accolement d'une boucle (= ansa) d'autres fibres nerveuses. Antagonistes : muscles agissant en sens inverse Apnée inspiratoire : arrêt de l'inspiration intra-utérine : dans l'utérus ataxie : maladie caractérisée par des mouvements non ciblés autocinétique : qui se déplace de lui-même bilatéral : des deux côtés C : nerfs cervicaux (= du cou) Carabus auratus : carabe doré Carcinus maenas : crabe de plage cataleptoïde : comme dans la "rigidité cireuse" (catalepsie), rester dans une position du corps avec une tension musculaire accrue cataphorèse : migration de substances solides en suspension dans l'eau dans un champ électrique Cécité de l'âme = agnosie optique : incapacité, due à une lésion de certaines parties du cerveau, de reconnaître et d'interpréter ce qui est vu tout en conservant la capacité visuelle. Cervelet : cerveau central Chémorécepteurs : terminaisons nerveuses sensibles pour la perception de stimuli chimiques Chorda thympani : branche du nerf facial qui conduit la sensation gustative des 2A antérieurs de la langue et fournit les muscles lisses des glandes salivaires de la bouche. chorée : contractions involontaires, incontrôlables et arythmiques rapides de muscles dans toutes les régions du corps, dues à une lésion du système extrapyramidal (voir là-bas) Contraction : contraction Contracture : réduction pathologique de la durée du muscle, crampe Réflexe coordonné : automatisme de mouvement coordonné contracture extrapyramidale : contraction des muscles provoquée par des mouvements extrapyramidaux. contracture hémiplégique : contraction des muscles d'une moitié du corps hémisection : demi-section Contraste de poils : si l'on pose sur une surface colorée une surface grise plus petite et que l'on recouvre le tout de papier mat ou d'un voile de soie en guise de "poil", la surface grise apparaît dans la couleur opposée si l'éclairage est approprié. corne postérieure : ensemble pair de cellules nerveuses afférentes dans la moelle épinière racine postérieure : points d'entrée des nerfs afférents dans la moelle épinière de fibres nerveuses se trouvant à l'extérieur des voies pyramidales. de la jambe et la face postérieure de la cuisse. Décérébration : suppression du cerveau par des processus pathologiques ou d'autres interventions, par exemple chirurgicales. Désafférentation : suppression neurochirurgicale des impulsions sensibles par une section correspondante des nerfs. Disparition transversale des zones rétiniennes : la répartition horizontale des zones rétiniennes symétriques dans les deux yeux, importante pour la vision spatiale. Dixippus (Carausius) morosus : criquet à tige Dysesthésie : sensation douloureuse Dytiscus marginalis : coléoptère du feu jaune, un coléoptère prédateur des eaux indigènes Echinodermes : échinodermes, par ex. étoile de mer ou oursin efférent : voie nerveuse s'éloignant du système nerveux central électroencéphalogramme : enregistrement de mesure et représentation graphique de l'activité électrophysiologique du cerveau électromyogramme : enregistrement par mesure et représentation graphique de l'activité musculaire. endosmotique : mode spécial d'échange de substances par diffusion, le solvant s'écoulant de l'extérieur dans une zone entourée d'une membrane semi-perméable. Exérèse du labyrinthe : ablation de l'oreille interne Extensor digitalis communis : le muscle qui étire tous les doigts ensemble Facial : voir nerf facial Fibre musculaire : fibre musculaire, sous-unité de la cellule musculaire striée. Fibrilles : fibres (ici des nerfs) Flagellés : Etres vivants unicellulaires avec des flagelles à rames (= flagelles) Flexor carpi radialis : muscle qui fléchit les os du carpe du côté du radius de l'avant-bras. Flexor carpi ulnaris : muscle qui fléchit les os du carpe du côté du coude de l'avant-bras. formation d'engrammes : la trace durable d'impressions sensorielles dans le cerveau, image-souvenir en sommeil. Fuseau musculaire : organe sensoriel situé dans le muscle, qui enregistre l'état de tension de ce dernier Gall, Franz Joseph (1758 - 1828) : médecin connu de l'époque de Goethe Ganglion dorsal : ganglion nerveux de la moelle épinière Ganglions du tronc : grands noyaux de cellules nerveuses ("masses grises") dans le diencéphale et le cerveau terminal pour la coordination des mouvements involontaires. Glossopharyngeus : voir nerf glossopharyngien hémianopsie : cécité d'un côté Hémiplégie de Brown-Séquard : due à une lésion de l'un des deux troncs de la moelle épinière. Hydrous : colombin d'eau Hypertonie : pression excessive, haute pression par exemple de la circulation sanguine Hypoglosse : voir nerf hypoglosse Hypothalamus : noyau central du diencéphale, situé entre le thalamus et l'hypophyse, qui exerce entre autres une activité glandulaire, c'est-à-dire sécrète des neurohormones. Hypotonie : dépression, basse pression, par ex. de la circulation sanguine Insertion : insertion (musculaire) Inspiration : prise d'air Inversion : retournement iodothyroglobuline : hormone thyroïdienne Irradiation : rayonnement d'une sensation dans la région entourant le point de stimulation lateralis : sur le côté les daphnies : puces d'eau, un groupe de petits crustacés d'eau douce Locuste migratoria : Criquet migrateur lors de la poursuite de plusieurs objets qui défilent successivement Macula : Macula lutea = tache jaune ; point de la vision la plus nette au centre de la rétine Moelle dorsale : partie postérieure de la moelle épinière Muscle droit interne (= médial) : muscle droit interne de l'œil]. Muscle droit supérieur : muscle droit supérieur de l'œil Muscle olliquus superior : muscle oblique supérieur de l'œil Muscle rectus externus (= lateralis) : muscle droit externe de l'œil Muscle rectus interior : muscle droit inférieur de l'œil Muscle semi-tendineux : muscle situé à l'arrière de la cuisse. Muscle tibial antérieur : muscle tibial antérieur Myopathie : maladie musculaire Nerf accessorius : nerf crânien moteur pour le muscle de l'inclinaison de la tête et le muscle de la capuche nerf cutané antébrachial : nerf cutané de l'avant-bras nerf facial : nerf facial principalement efférent nerf gastrocnémien : nerf de la peau et des muscles du mollet nerf glossopharyngien : nerf de la langue et du pharynx nerf hypogastrique : nerf qui transmet les sensations de la région latérale de la hanche et du pubis Nerf hypoglosse : Nerf sous-glottique nerf iléo-inguinal : nerf conduisant les sensations des lèvres de la femme et du scrotum de l'homme nerf infraorbitaire : nerf de la mâchoire supérieure, de la lèvre supérieure et de la partie inférieure du nez Nerf laryngé supérieur : nerf laryngé supérieur Nerf lingual : nerf assurant la sensibilité des amygdales palatines, du plancher buccal, des gencives et des 4 antérieurs de la langue. Nerf médian : nerf des muscles et de la peau d'une région de l'avant-bras, de la main creuse et de l'éminence thénar. Nerf musculo-cutané : nerf innervant le biceps du bras et la face externe de l'avant-bras. Nerf musculo-spiral : nerf pour la peau et les muscles d'une région spécifique de l'avant-bras nerf péronier : nerf pour la peau et la musculature de la face externe de la jambe et du dos du pied. Essentiel pour soulever la pointe du pied pendant la marche. Nerf phrénique : approvisionne le péricarde, la plèvre et le diaphragme. Nerf récurrent : approvisionne les muscles du larynx, responsables de la formation de la voix. nerf sciatique : cordon nerveux qui innerve le bord externe du pied, la face externe de la jambe et la partie supérieure de la jambe. Nerf sympathique : partie du système nerveux involontaire qui commande des processus corporels inconscients permettant une libération d'énergie (p. ex. en cas de fuite, de stress, de peur ou de froid). nerf tibial : nerf de la peau et des muscles dans la région du tibia nerf trijumeau : nerf facial principalement afférent avec alimentation efférente des muscles masticateurs Nerf ulnaire : nerf pour la peau et les muscles dans la région du cubitus de l'avant-bras Nerf vague : partie du système nerveux involontaire pour la commande de processus corporels inconscients qui permettent une fixation de l'énergie (par ex. par le sommeil, la digestion) ; antagoniste du nerf sympathique (voir là-bas) Neurofibrilles : structures fibreuses à l'intérieur des cellules nerveuses neurone : cellule nerveuse Neurotisation, névrosé : Production de réactions anormales et continues à l'expérience. Névrose explosive : comportement erroné explosif, déclenché par l'âme Expiration : action d'expirer Nucleus ruber = noyau rouge : zone importante du tronc cérébral pour la régulation de la posture et des mouvements de marche. Nystagmus : mouvement lent des yeux dans le sens de la rotation, suivi d'un retour saccadé. nystagmus rotatif optocinétique : mouvement de retour saccadé des yeux ontogénétique : qui concerne le développement d'un organisme individuel Paralysie radiale : paralysie des muscles extenseurs et perte de la sensibilité de l'avant-bras en cas d'interruption de la conduction du nerf radial. Parésie : paralysie partielle Parkinsonisme : syndrome caractérisé par des symptômes physiques et psychiques d'origine extrapyramidale et végétative ; déclenché par l'atrophie d'une zone spécifique du cerveau (globus pallidus et substantia nigra), principaux symptômes : akinésie, rigidité, tremblement, dépression Périmétrie : détermination du champ visuel Périmysium externe : enveloppe des muscles squelettiques Périmysium interne : enveloppe des fibres musculaires squelettiques Phalanges des membres : extrémités osseuses des membres (doigts et orteils) phylogénétique : concernant la phylogénie Planaires : voir turbellariés Plasmodium = syncytium : voir là-bas Plexus brachial : plexus nerveux du bras, à partir duquel s'effectue la division en différents troncs nerveux pour le bras. Plexus mucosus : plexus nerveux dans la muqueuse intestinale pour la régulation du péristaltisme et de la sécrétion glandulaire. Poliomyélite antérieure = poliomyélite : dégénérescence inflammatoire de la substance grise des cornes. Proprioceptivité : perception propre proximale : dirigée vers le tronc réflexe conditionnel : un réflexe primaire inné est secondairement lié par apprentissage à un stimulus étranger spécifique. Celui-ci peut alors également déclencher le réflexe. Réflexe d'étirement ipsilatéral : automatisme de mouvement équilatéral conduisant à l'étirement réflexe de Babinski : réflexe du gros orteil, extension paresseuse après un passage vigoureux sur le bord latéral du pied, qui indique une lésion de la voie pyramidale (voir là-bas). Réflexe propre : automatisme de mouvement inconscient dans lequel l'organe de stimulation et l'organe de succès sont identiques. Région cortico-pyramidale : région et connexion du cerveau avec les voies pyramidales (voir là-bas). Région extrapyramidale-cérébrale : lien entre le système extrapyramidal (voir là-bas) et le cervelet. réseau de fibrilles : réseau de fibres microscopiques Réticulum terminal : réticulum terminal Rigidité : rigidité Rigidité : tension de base accrue de la musculature squelettique avec rigidité typique lors de mouvements passifs. Segments lombaires : Sections de la moelle épinière dans la région lombaire Spasmes : contractions, crampes, éventuellement rythmiques. substance des cornes de Vörder (neurones efférents) de la moelle épinière. substances de type mon dans le sang à partir des cellules nerveuses. Sympathique : voir nerf sympathique. synapse : la fente de contact entre deux cellules nerveuses synchrone : simultané syncytium : structure multinucléée sans frontières cellulaires système cellulaire neurohumoral : le système cellulaire qui produit des hormones tissulaires ou des hormones hor-. Système extrapyramidal : partie du système nerveux central qui, contrairement au système pyramidal (voir là-bas), régule la posture et les mouvements involontaires du corps. Système pyramidal : partie du système nerveux qui, contrairement au système extrapyramidal (voir là-bas), régule la posture et les mouvements volontaires du corps. Tabes dorsalis : dégénérescence de la moelle épinière, vertige médullaire. Forme tardive de la syphilis Tache aveugle : point d'entrée du nerf optique dans la rétine de l'œil Système du canal semi-circulaire : organe sensoriel du mouvement de rotation dans l'oreille interne. Tachystoscopie : observation en lumière rasante, par ex. en lumière scintillante Ténotomie : section d'un tendon téléologique : approprié Th : nerfs thoraciques ( = thorax) Tissu chromaffine : tissu pouvant être coloré par des sels de chrome ; produit l'adrénaline dans la glande surrénale. Tonus : tension Tractus rubrospinal : voie nerveuse extrapyramidale du nucleus ruber, l'un des ganglions du tronc (voir là-bas), vers les cornes antérieures de la moelle épinière. trigéminal : voir nerf trijumeau triton : triton trophique : alimentaire Turbellariés : turbellariés, groupe de vers plats vivant librement dans l'eau (Plathelminthes) vasomoteurs : Nerfs des vaisseaux sanguins du système nerveux végétatif Organe vestibulaire : l'organe de l'équilibre dans l'oreille interne Cervelet : cervelet Vertèbre sacrée : vertèbre du sacrum voies afférentes : voies nerveuses menant au système nerveux central Voies pyramidales : ensemble des voies nerveuses allant du cortex cérébral à la moelle épinière. Indispensable pour les mouvements volontaires en tant que partie du système pyramidal (voir là-bas). Petite biographie de Gerhard Kienle par Peter Selg Gerhard Kienle a été sans conteste l’un des anthroposophes les plus efficaces et les plus performants après la Seconde Guerre mondiale. Il a réussi, dans le cadre d’un travail de pionnier sans précédent, à fonder la première maison anthroposophique des étudiants à Tübingen, une grande clinique anthroposophique dans la région de la Ruhr ainsi que la création d’une université privée reconnue par l’État — sous le signe d’une vie de l’esprit libre et d’un traitement futur de questions scientifiques et sociales centrales. En outre, Kienle a réussi à mener une activité scientifique — avec un doctorat et une habilitation sur des questions de recherche originales de Steiner —, à faire évoluer la loi sur les médicaments en Allemagne en faveur d’un pluralisme méthodologique et à acquérir une grande réputation scientifique et publique. Bien que méconnu de son vivant par de nombreux anthroposophes, il travailla dans une fidélité contraignante aux préoccupations fondamentales de Rudolf Steiner, s’engagea sans compromis et de toute son existence pour celles-ci et créa de grands espaces de développement social dans un style constamment spirituel et avec une attention vigilante, résolument tournée vers l’avenir, pour les forces de l’histoire contemporaine. « Vouloir être efficace, c’est se préparer à être capable de répondre aux exigences du destin. Cela exige de saisir et de se placer dans le moment présent. L’efficacité active exige une écoute intensive du langage du destin, un engagement total ». (Lettre de Gerhard Kienle à Gisela Hörtreiter, 17.6.1956) Gerhard Kienle est né le 22 novembre 1923 à Madrid, où son père, qui faisait partie du service diplomatique du ministère des Affaires étrangères, travaillait temporairement. En 1926, la famille est retournée à Berlin, mais l’enfance et l’adolescence de Gerhard Kienle ont été marquées par des séjours répétés et prolongés à l’étranger pour se former l’horizon grâce à l’activité professionnelle de son père, par exemple en Estonie (1935) et au Guatemala (1935/36). Wilhelm Kienle et son épouse Erika, née Krüger, qui travaillait de manière engagée comme assistante sociale, étaient des humains cherchant spirituellement, largement dépourvus de patrie qui se sont affiliés à différents groupes religieux, dont les francs-maçons, les rosicruciens et les mazdaniens. Kienle lui-même, à l’âge de 16 ans, se lia par des voies indépendantes à la communauté des chrétiens — en particulier au prêtre berlinois Otto Palmer —, tout en restant dans un premier temps dans une relation vigilante et pondérée avec l’anthroposophie, dont il considérait les résultats de travail comme des hypothèses scientifiques et commençait à les vérifier. Dans une attitude distanciée vis-à-vis du régime national-socialiste, il commença en 1941 à Berlin ses études de médecine à l’Académie médicale militaire, fonda pour la première fois en 1943 à Würzburg, dans la clandestinité, un groupe anthroposophique d’étudiants qu’il voulait préparer à la fondation d’une université libre après la fin de la guerre mondiale, et fut blessé à l’automne 1944 près d’Arnheim dans son activité de médecin auxiliaire. Alors qu’il était encore à l’hôpital, il développa des plans pour la création d’un nouveau type d’hôpital, social et fondé sur l’anthroposophie, afin de pouvoir préserver la liberté de la médecine dans le sens de la possibilité de destin d’un art de guérir déterminé au Je du malade, face à toutes les influences étatiques et socio-administratives. Après la fin de la guerre, il forma à nouveau un groupe anthroposophique d’étudiants à Tübingen, où il poursuivit ses études de médecine à partir d’octobre 1945, mais cette fois avec un discours public et rayonnant. Avec quelques amis d’études anthroposophiques (comme Siegfried Gussmann et Oleg Selawry entre autres) et avec le soutien de Diether Lauenstein, un travail strictement méthodique sur l’œuvre de Steiner se développa, complété par des groupes de travail spécialisés et des conférences publiques au sein de l’université et dans le cadre du groupe. Avec Diether Lauenstein, Kienle s’est attaché à mettre en place des institutions culturelles efficaces dans le sens d’une vie de l’esprit libre. Tous deux ont tenté de préparer le terrain méthodologique et de rassembler les gens à Tübingen, en vue de la fondation ultérieure d’un hôpital et d’une université et d’une prise d’influence sur le développement académique et culturel de l’Occident, en particulier sur un futur ordre social de la médecine et de la société. Kienle considérait les Semaines universitaires de Stuttgart (à partir de 1947) comme une « répétition générale pour la possibilité de réaliser une université indépendante de l’État, portée par l’anthroposophie » — en 1950, lors d’une assemblée de la Société nationale allemande, il déposa pour la première fois une motion pour l’union de tous les séminaires anthroposophiques existant jusqu’alors à Stuttgart (Communauté chrétienne, École Waldorf, Année d’études libres) comme étape préalable à la fondation d’une université indépendante. Ses projets ne rencontrèrent cependant aucun écho au sein des instances dirigeantes de l’époque. — Parallèlement, Kienle se lança dans un travail scientifique et, après son doctorat en 1948, il poursuivit la question des « nerfs moteurs » au sein de l’Institut de physiologie de l’université de Tübingen, sur laquelle il élabora de nombreux manuscrits en s’appuyant sur toute la littérature scientifique (entre autres « Die Grundfragen der Nervenphysiologie », 1950). La réception de ces travaux par le corps médical anthroposophique a également été durablement décevante pour Kienle. Après des travaux cliniques à Tübingen et des études de recherche homéopathique à Stuttgart, Kienle partit début 1951 pour deux ans et demi en Suisse, où il s’occupa médicalement de deux foyers de pédagogie curative, fit des recherches neurophysiologiques à l’université de Zurich et prépara en même temps la fondation d’un futur « hôpital communautaire » anthroposophique — par des séminaires de formation spéciaux (entre autres avec Herbert Sieweke et Ernst Marti), des élaborations théoriques et l’examen des premières reprises de bâtiments et options de construction. En automne 1953, Kienle commença une formation universitaire de médecin spécialiste auprès du psychiatre renommé Ernst Kretschmer à Tübingen et rédigea dans les années qui suivirent ses premières publications remarquables sur un grand nombre de sujets psychiatriques, neurologiques, et anthropologiques-anthroposophiques. En 1957, il conclut une alliance de vie avec le médecin anthroposophe Gisela Hörtreiter. À la fin des années 50, Kienle commença, parallèlement à son activité à la clinique universitaire, ses études extensives de neurophysiologie, de mathématiques et de statistiques, qui mirent en évidence la structure non-euclidienne de l’espace visuel humain et qui devaient aboutir en 1966 à son habilitation et à la publication d’un livre (qui suivit), ainsi qu’aux travaux préparatoires communs avec Diether Lauenstein pour un premier foyer d’étudiants anthroposophique, qui ouvrit finalement ses portes en 1963 sous le nom de Johann Gottlieb Fichte-Haus et déploya une activité extrêmement fructueuse. Parallèlement, Kienle s’est penché sur d’importants problèmes de sa spécialité médicale et a non seulement rédigé une monographie internationalement reconnue sur le traitement d’urgence en neurologie et en psychiatrie, mais il a également participé aux discussions de l’époque sur l’organisation future d’un hôpital moderne avec un autre ouvrage ; en outre, au début des années 60, il s’est penché pour la première fois de manière plus intensive sur les développements à venir de la législation sur les médicaments et de la politique de santé et a plaidé au sein de l’Association des médecins anthroposophes (il était membre du comité directeur élargi) pour la création de propres commissions sur les médicaments afin d’élaborer des méthodes spécifiques de preuve de l’efficacité des médications anthroposophiques-homéopathiques, mais à nouveau sans aucun écho. Kienle a toujours défendu l’idée que la tâche des anthroposophes était de s’occuper de manière existentielle des problèmes mondiaux actuels, de ne pas « faire du prosélytisme dans les solutions », mais de montrer, d’imaginer et d’élaborer de nouvelles voies en luttant sur les questions de fond — et d’acquérir ainsi un respect et une reconnaissance professionnels dans le monde. Bien qu’il ait travaillé toute sa vie en toute loyauté avec des institutions anthroposophiques centrales (comme l’École supérieure de Dornach, l’École supérieure de la communauté chrétienne de Stuttgart ou la Société nationale allemande, dont il était membre du conseil d’administration) — des institutions qu’il considérait comme des facteurs importants de civilisation ou qu’il voulait développer —, Kienle trouvait que leurs activités étaient la plupart du temps beaucoup trop circonscrites, trop égocentriques et passaient à côté des véritables intentions de Rudolf Steiner. Kienle voulait que l’anthroposophie soit offensive, qu’elle contribue à façonner la réalité sociale dans son ensemble et qu’elle en soit coresponsable, et qu’elle s’engage en même temps dans le discours scientifique, qu’il avait reconnu comme un objectif central de la vie de Rudolf Steiner. Mais il s’est souvent retrouvé seul dans cette démarche. De 1964 à 1969, Gerhard Kienle travailla comme chef de clinique dans l’un des services de neurologie les plus modernes de Francfort, devint privat-docent à l’université Johann Wolfgang von Goethe, organisa des symposiums scientifiques et donna des conférences ; en outre, il fit, avec son collègue médecin Anselm Basold, l’essentiel du travail de préparation de l’hôpital communautaire de Herdecke, qui fut finalement mis en service en novembre 1969 — après presque 18 ans d’efforts de la part de Kienle et la perte de tous les anciens compagnons d’armes — et bénéficia d’une grande participation publique. Herdecke" était le premier hôpital de soins généraux anthroposophiques. « Hôpital général » avec des soins aigus et près de 200 lits ; son ouverture a eu lieu à une époque où la société tenait un discours intense sur la liberté de l’homme, le démantèlement des systèmes sociaux autoritaires et la « maturité » du patient au sein d’une médecine qui était majoritairement vécue comme niant l’individualité et centrée sur la technique. Kienle lui-même était le porte-parole du collège des médecins dirigeants et travaillait sous la responsabilité principale du service de neurologie de l’établissement — il était le Spiritus Rector incontesté du lieu, également le centre social de la grande fondation. Moins directif que convaincant, il se tenait au centre, connaissait les individus et favorisait leur développement avec une grande force d’encouragement et de lumière. En 1970, une école d’infirmières associée a déjà pu être ouverte, dans le sens de l’intention de Kienle d’ériger des jalons modèles et efficaces pour l’humanisation et donc l’individualisation de la médecine — en direction d’une christianisation de l’art de guérir et à l’aide de l’anthroposophie. La même année, Kienle s’engagea résolument dans les débats qui reprirent au sujet d’une réforme de la loi allemande sur les médicaments qui, à cette époque, devait être transformée, suite aux premières « catastrophes médicamenteuses » (thalidomide, ménocil) et à la pression internationale, en un instrument de sélection de substances « efficaces » sur la base de procédés scientifiques dépourvus de sujet (essai en double aveugle), avec des tests obligatoires sur les animaux et l’autorisation correspondante d’une autorité de surveillance étatique (Office fédéral de la santé). Malgré de nombreuses mises en garde mesurées et centrées sur la préservation des intérêts propres, Kienle n’a pas mené la discussion dans le but d’obtenir des dérogations pour certains médicaments, mais de remettre en question, voire de réfuter, les paradigmes scientifiques des systèmes de contrôle formels et dirigistes largement propagés et de faire évoluer la médecine humaine dans un esprit de critique des connaissances et en tenant compte de l’individualité de l’être humain. Kienle a poursuivi ce combat scientifique sans interruption jusqu’en 1983, avec le soutien de quelques collaborateurs seulement (comme Rainer Burkhardt en particulier), mais dans l’ensemble avec un succès remarquable. C’est notamment grâce aux expertises et aux arguments scientifiques de Kienle que la loi sur les médicaments de 1976 a été rédigée dans un esprit de pluralisme des méthodes, ce qui a relativisé le droit de domination de la médecine universitaire, mais c’est également à ses interventions que l’on doit d’autres évolutions importantes (par exemple en ce qui concerne l’ordonnancement de l’admission des médecins ou la loi sur les soins infirmiers). A Bonn, Kienle était à l’époque une figure reconnue depuis longtemps et un partenaire de discussion apprécié, bien que redouté, dans les commissions spécialisées et les symposiums. Sa recherche de la vérité, sa capacité de jugement individuel et son engagement suscitaient le respect, même dans les rangs de ses adversaires. À partir de 1976, Kienle a tenté, en collaboration avec Diether Lauenstein, mais sans succès, de mettre en place un système d’évaluation de la qualité, également avec Herbert Hensel et Karl Ernst Schaefer, de poursuivre les discussions de fond entamées avec le positivisme dominant dans un cadre plus large et de mettre en œuvre la fondation d’une université prévue depuis longtemps. Après un premier symposium couronné de succès, Kienle réussit à créer une « Fondation Académie libre européenne des sciences (FEAW) » en été 1976 et à réunir plus de 60 professeurs d’université internationaux ayant des préoccupations anthroposophiques et anthropologiques ; dès l’automne de la même année, il établit un cursus réformé de médecine à Herdecke et entra ainsi dans les préparatifs proches d’une réelle fondation d’université. Avec une petite équipe de collaborateurs scientifiques, Kienle, Lauenstein et Hensel firent avancer le travail conceptuel, tandis que le médecin Konrad Schily encouragea durablement les processus et les décisions politiques. Malgré des controverses internes à l’anthroposophie, à nouveau épuisantes, la participation publique à cette université modèle fut à nouveau élevée et décisive — en été 1982, l’État donna son autorisation. — Mais lorsque l’université ouvrit ses portes fin avril 1983, Gerhard Kienle, dont c’était l’œuvre de toute une vie, fut gravement malade dans le service de soins intensifs de l’hôpital de sa communauté, « enserré dans tout l’arsenal de la médecine moderne du XXe siècle (Dumke) qu’il avait pris sur lui de surmonter ». C’est là que Kienle s’est éteint le 2 juin 1983, jour de la Fête-Dieu, après une vie de dévouement total et de sacrifice de soi. Peter Selg
original allemand avec
photographies
https://biographien.kulturimpuls.org/detail.php?&id=175 Œuvres : Das Gemeinschaftskrankenhaus, Tübingen 1961 ; Notfalltherapie neurologischer und psychiatrischer Erkrankungen, Stuttgart 1964, 31978 ; en tant qu’éditeur : Hydrodynamik, Elektrolyt-und Säure-Basen-Haushalt im Liquor und Nervensystem, Stuttgart 1967 ; Die optical Wahrnehmungsstörungen und die nicht-euklidische Struktur des Sehraums, Stuttgart 1968 ; Arzneimittelsicherheit und Gesellschaft. Eine kritische Untersuchung, Stuttgart 1974 ; avec Burkhardt, R. : Die Zulassung von Arzneimitteln und der Widerruf von Zulassungen nach dem Arzneimittelgesetz von 1976, Stuttgart 1982 et Der Wirksamkeitsnachweis für Arzneimittel. Analyse einer Illusion, Stuttgart 1983 ; Die ungeschriebene Philosophie Jesu, Stuttgart 1983 ; Christentum und Medizin, Stuttgart 1986 ; contributions dans des ouvrages collectifs ; traductions en différentes langues ; nombreuses contributions dans des revues spécialisées et BeH, d’autres dans Arb, BaS, DD, MaD, Wa, WKÄ. Littérature : Lauenstein, D. : Gerhard Kienle, dans : MaD 1983, M— 145, également dans : Selg, P. [éd.] : Anthroposophische Ärzte, Dornach 2000 ; Hemmer, W. : Gerhard Kienle. Il a nagé à contre-courant, in : Medicale Tribune, 14.10.1983 ; Schily, K. : Dr. med. habil. Gerhard Kienle, in : N 1983, M— 33 ; Büttner, G. : Im Gedenken an Gerhard Kienle, in : BeH 1983, M- 5 ; Gussmann, M. : Gerhard Kienle, in : DD 1983, M- 7/8 ; Worel, A. : Gerhard Kienle - ein Streiter für das freie Geistesleben, in : BaS 1983, M- 15 ; Wittich, J. : Zum Tod von Gerhard Kienle, in : I3 1983, M- 7/8 ; Dumke, K. : Gerhard Kienle, in : Jahrbuch der Anthroposophischen Gesellschaft in Deutschland 1984 ; Selg, P. : Gerhard Kienle und die Humanisierung der Medizin, in : Mst 1999, M- 5 ; ders : Anfänge anthroposophischer Heilkunst, Dornach 2000 ; ders : Spatenstiche des Willens tun, in : G 2001, M-9 ; ders : Gerhard Kienle. Leben und Werk, vol. I/II, Dornach 2003. Abréviations : voir www.kulturimpuls.org Copyright : le texte et les images sont protégés par le droit d’auteur. Reproduction sous quelque forme que ce soit uniquement après autorisation écrite du Centre de recherche Kulturimpuls, Dornach Centre de recherche Kulturimpuls Biographies Documentation www.kulturimpuls.org |